爬行动物的演化历史
爬行动物的演化历史
在最早的爬行动物出现后不久,出现了两个演化支,一个是无孔亚纲。无孔亚纲拥有坚硬的头颅骨,没有颞颥孔,仅有与鼻孔、眼睛、脊椎相对应的洞孔。乌龟被认为是目前仅存的无孔动物,因为它们拥有相同的头颅骨特征;但最近有些科学家认为乌龟是返祖遗传到原始的状态,以增加它们的保护能力。另一群演化支是双孔亚纲,头颅骨上有两个颞颥孔,位于眼睛后方。双弓动物进一步分化为两个支系:鳞龙类包含现代蜥蜴、蛇、喙头蜥,可能还有中生代的已灭绝海生爬行动物;主龙类包含现代鳄鱼与鸟类,以及已灭绝的翼龙目与恐龙。
而最早期、具坚硬头颅骨的羊膜动物也演化出另一独立的演化支,称为单孔亚纲。单弓动物的眼睛后方有一对窝孔,可减轻头颅骨重量,并提供颌部肌肉附着点。单孔亚纲最后演化为哺乳类,因此被称为似哺乳爬行动物。单孔亚纲过去为蜥形纲的一个亚纲,但目前为个别的合弓纲。
现代的爬行动物栖息于每个大陆,除了南极洲以外,但它们主要分布于热带与副热带地区。现存的爬行动物,体型最大的是咸水鳄,可达7米以上,最小的是侏儒壁虎,只有1.6厘米长。除了少数的龟鳖目以外,所有的爬行动物都覆盖者鳞片。
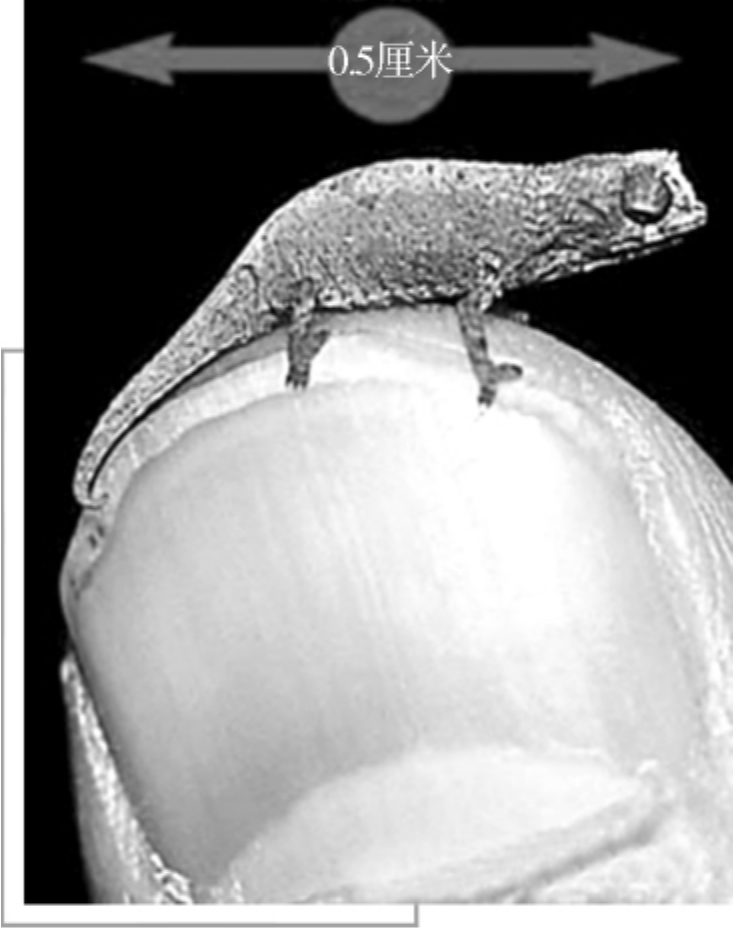
侏儒壁虎
虽然所有的细胞在新陈代谢时都会产生热量,大部分的爬行动物不能产生足够的热量以保持体温,因此被称为冷血动物或变温动物(棱皮龟可能是个例外)。爬行动物依靠环境来吸收或散发内部的热量,例如在向阳处或阴暗处之间移动,或借由循环系统将温暖血液流动至身体内部,将较冷血液流动至身体表层。大部份生存于天然栖息地的爬行动物,可将身体内部的体温维持在相当狭窄的变化范围内。不像两栖类,爬行动物的表皮厚,因此不需要栖息在水边,吸取水份。由于体温调节方面的关系,爬行动物可以较少的食物维生。温血动物通常以较快速度移动,一种攻击性蜥蜴、蛇、或鳄鱼的移动速度较快。
大部分的爬行动物是卵生动物,它们的胚胎由羊膜所包覆。然而,许多有鳞目能够直接生下后代,例如卵胎生或胎生的方式。大部分的胎生爬行动物借由不同形式的胎盘给与胎儿养分,类似哺乳类的方式。它们通常提供刚出生幼体适当的初步照顾。
循环系统
大部分的现存爬行动物具有闭合的循环系统,它们具有三腔室心脏,由两个心房与一个心室所构成,心室的分割方式并不一致。它们通常只有一对大主动脉。当它们的血液流经三腔室心脏时,含氧血与缺氧血只有少量混合。但是,血液可改变流通方式,缺氧血可流向身体,含氧血可流向肺脏,使爬行动物的体温调节更有效率,尤其是水生物种。
鳄鱼具有四腔室心脏,可以在水中以三腔室心脏运作。某些蛇类与蜥蜴(例如巨蜥与蟒蛇),具有三腔室心脏,但可以四腔室心脏方式运作。因为它们心脏的皮瓣可在心动周期时,随者扩张、收缩隔开心室。某些喙头蜥可借由皮瓣,产生类似哺乳动物与鸟类的心脏运动。
呼吸系统
所有的爬行动物都用肺脏呼吸。水生乌龟发展出具渗透性的皮肤,某些爬行动物可用泄殖腔来增加气体交换的面积。即使具有这些构造,它们仍需要肺脏来完成呼吸作用。在各个主要的爬虫类动物种类之间,完成肺部换气的方式并不相同。有鳞目爬虫类的肺部几乎只借由轴肌换气,而这个肌肉也同时用于运动。因为这个限制,大部分的有鳞目爬虫类必须在激烈的移动中停止呼吸。但是,有些爬虫类找到了解决方法。巨蜥以及一些其他的蜥蜴物种使用一种使用到喉咙肌肉的方法来补足原本的“轴呼吸”。这使得这些动物可以在激烈运动中为肺部完全充气,所以可以持续进行有氧呼吸很长一段时间。据了解双领蜥具有原生横隔膜可将胸腔和腹腔分开来。即使此膜不能动,它还是能借着将内脏的重量和肺分开而增加肺的吸气量。鳄鱼其实拥有和哺乳类动物相似的有肌肉横隔膜。不同的是,鳄鱼的横隔膜将耻骨拉回(部分的骨盆,在鳄鱼中能移动),从而使肝下降,以腾出空间使肺能胀大。这种横隔膜式的机制被称作“肝活塞”。
关于海龟和陆龟如何呼吸已经被研究了很多次。迄今为止,只有少数物种已经被深入研究到足以了解乌龟如何呼吸。其结果表明,海龟及陆龟分别找到了不同的方法来解决这个问题。但问题是,大部分的龟壳是僵硬的,且不能像其他羊膜动物使用的膨胀和收缩来为肺部换气。一些乌龟,如印度鳖(lissemys punctata)有一层肌肉包覆著肺部。当它收缩时,乌龟可以呼气;在静止时,乌龟可以收回四肢进到体腔内,并强制使空气排出肺部的。而当乌龟伸出其四肢时肺部内的压力会减少,乌龟因此可以吸进空气。乌龟的肺连接到壳内的上方,而肺的底部附着在其余内脏(由结缔组织连接)。借由一系列特殊的肌肉(大约相当于一个横隔膜),因为许多这些肌肉的附着点都在他们的前肢上(事实上,很多的肌肉在收缩时扩张到四肢的袋子里),乌龟能够推动他们的内脏向上或向下,造成有效的呼吸。
现今已研究了3个物种在运动期间的呼吸,并且显示了它们之间不同的模式。当成年雌性绿蠵龟沿着要产卵的海滩爬行时并不呼吸。他们在陆上运动时屏住呼吸,只有在停顿休息时呼吸。北美的箱龟运动期间还是继续呼吸,而且其换气的周期并不与肢体动作相关。它们可能是利用其腹部肌肉在运动中呼吸。最后被研究的物种是红耳龟,它们也在运动中呼吸,但他们在运动中的呼吸比在运动间停顿时的呼吸还少,显示肢体运动和呼吸器具之间可能有机械干扰。箱龟也被观察到在完全密封的壳内呼吸。

绿蠵龟
大部分的爬行动物缺乏次生颚,所以当它们进食时,无法同时呼吸。鳄鱼已发展出骨质次生颚,使它们可在半隐没至水中时持续呼吸,并防止嘴中的猎物挣扎时,伤及脑部。石龙子科也演化出骨质次生颚。
排泄系统
爬行动物的排泄系统主要借由两颗肾脏。双孔动物所排泄的主要含氮废物是尿酸;而乌龟主要排泄尿素,类似哺乳类。不像哺乳类与鸟类的肾脏,爬行动物的肾脏不能够制造尿液,尿液可以储藏更多的身体废物。这是因为它们缺乏鸟类与哺乳类所拥有的肾元,称为亨利氏环。因此许多爬行动物使用它们的结肠与排泄腔来再度吸收水分。有些爬行动物甚至可利用膀胱来取得水分。有些爬行动物的鼻部与舌部具有盐腺,可以储存额外的盐分。
骨骼系统
爬行动物的骨骼系统大多数由硬骨组成,骨骼的骨化程度高,很少保留软骨部分。
神经系统
爬行动物与两栖类的脑部具有相同的基本部分,但它们的大脑与小脑稍大。爬行动物的感觉器官多发展良好,除了少部份物种,例如蛇缺乏外耳,但仍具有中耳与内耳。爬行动物具有12对脑神经。
繁 衍
除了陆龟与海龟以外,大部分的雄性爬行动物具有成对的管状性器官,称为半阴茎。陆龟与海龟则具有单一阴茎。所有的龟鳖目皆为卵生动物,而某些蜥蜴与蛇是卵胎生或胎生动物。爬行动物借由泄殖腔来交配、繁衍;泄殖腔位于尾巴基部,可用来排泄与繁殖。

乌龟及其龟蛋
对于某些有鳞目物种而言,雌性个体若达到一定的数量,它们便能自行复制出单性染色体。这种无性繁殖方式称为孤雌生殖,目前已有6科蜥蜴,以及一种蛇,被确认具有这种无性繁殖能力,例如:数种壁虎、美洲蜥蜴科、以及蜥蜴科。被封闭的科莫多龙(巨蜥科)也具有孤雌生殖能力。除了以上物种以外,变色龙、飞蜥科、夜蜥蜴科、以及嵴尾蜥科,也被推测具有孤雌生殖能力。
爬行动物的蛋,外部是钙质蛋壳或皮革,覆盖者内部的羊膜、羊膜囊、以及尿囊。
最早的爬行动物——林蜥
林蜥(学名: Hylonomus)是种早期的爬虫类,它生存于3.15亿年前的石炭纪。到2006年为止,它是目前所能确认最早的爬虫类;西洛仙蜥更为古老,但应为两栖类,但目前被认为较接近于两栖类,而非羊膜动物。它大约20厘米长(包括尾巴),而且外表应该相当接近现代蜥蜴,它拥有锐利的小型牙齿,应该以千足虫与早期昆虫为生。林蜥是后来所有爬虫类的祖先。油页岩蜥与中龙都为最早期的爬行动物之一。最早期的爬行动物被发现于石炭纪的沼泽森林,但体型小于同时期的迷齿螈类。石炭纪末期的小型冰河期,使得早期爬行动物有机会成长至较大的体型,例如基龙与异齿龙。
科学家在加拿大找到了当时巨大的封印木的化石,并惊喜地发现,在植物化石的空树干中,还躲藏着非常原始的爬行动物。人们给这种动物起名叫林蜥,它们到底是为了避难而躲在树洞中,还是就居住在其中,人们不得而知。从林蜥的骨骼化石观察,它的头骨很高,口中具有迷宫样式的迷齿,这些都是两栖动物的特点;而林蜥的脊椎骨则与其后出现的爬行动物相似,这个长有两栖动物的头和爬行动物的身子的怪家伙,被定为爬行动物的老祖宗。