交换的机制
如果像其他证据清楚地显示的那样,染色体是基因的携带者,如果基因在同对染色体中可能发生交换,那么迟早有希望找出这一交换发生的某种机制。
在遗传学发现交换作用的若干年之前,染色体的接合过程以及成熟生殖细胞中染色体数目的减少都已经完全确定了。已经证实的是,接合时同一对染色体就是互相接合的那两条。换句话说,接合并不像之前对接合过程所做的推测那样是随机的,接合总是发生在来自父方与母方的特殊染色体之间。
我们现在可以补充这样一个事实,即接合之所以发生是因为一对(两条)染色体是相似的,而不是因为它们分别来自雄体与雌体。这可以通过两个方面来证明:在雌雄同体类型中,尽管是自体受精,每一对的两条染色体都来自同一个个体,但同样的接合依然会发生:其次,在个别例子中,同对的染色体虽然来自同一个卵子,但由于交换发生,所以仍然可以假定它们会接合。
相似染色体接合的细胞学证据,对交换发生的机制进行了初步解释,很显然,如果每对的两条染色体整条并列,就像成对基因两两并列一样,在这些位置可以引起相对两段染色体之间有秩序的交换。当然,不能就此认为,染色体并列排列,交换就必然发生。事实上,一项关于连锁群内交换的研究,例如果蝇性连锁基因群(在这一连锁群中有足够的基因数量,提供关于连锁群内发生变化的充分证据),表明有43.5%的卵子内的成对染色体没有发生交换,同样的证据显示在43%的卵子中发生了一次交换,发生两次交换(双交换)的占13%,发生三次交换的占0.5%。在雄性果蝇中,一次交换也没有发生。
1909年,Janssens发表了一份关于他称其为“染色体交叉型”的详尽报告。我们这里不谈Janssens研究成果的详细内容,只提他相信他能够提供证据来证明,在接合的成对的2条染色体之间存在着整段交换,这种交换可以追溯到接合的2条染色体在早期的互相缠绕(图3-6)。
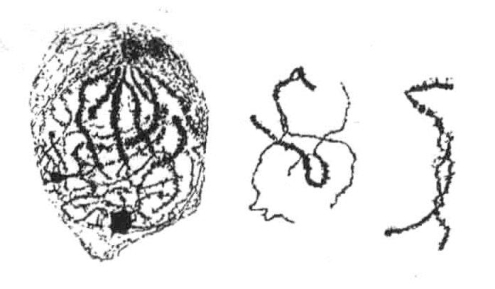
图3-6 Batrachoseps(蜥尾螈属)染色体的接合
Batrachoseps染色体的接合,中间图片中的两条染色体中的一条有两条细丝相互缠绕(根据Janssens)
可惜的是,在成熟分裂中,与其他任何阶段相比,染色体缠绕时期引起了更多的争论。就这一案例的实质来说,即便承认染色体缠绕的发生,实际上也可能证实它能够引起遗传学证据所要求的那种交换。
染色体彼此缠绕的图片已发表了很多。在某些方面,这一证据并不让人信服。例如,在最常见和最确定的有缠绕明显存在的阶段,就是当接合成对的染色体变短准备进入纺锤体的赤道面时(图3-7)。通常把这一阶段的染色体的缠绕解释为与2条接合染色体的变短存在某种联系。在这些图片里,没有迹象表明会引发交换。虽然此类案例中有一些是早期缠绕的结果,不过螺旋状态的持续存在更显示出没有发生交换,因为交换会解开缠绕。
如果看一下早期阶段的图片,我们发现有大量例子证明,细丝(细丝时期)似乎是相互缠绕的,见图3-8(b),不过这种解释经常引起质疑。如何判定二者在各个接合点上谁处于上方,谁处于下方,事实上是很困难的。更困难的是,细丝只有在凝固状态下才可以着色,以便能在显微镜下观察。(https://www.daowen.com)
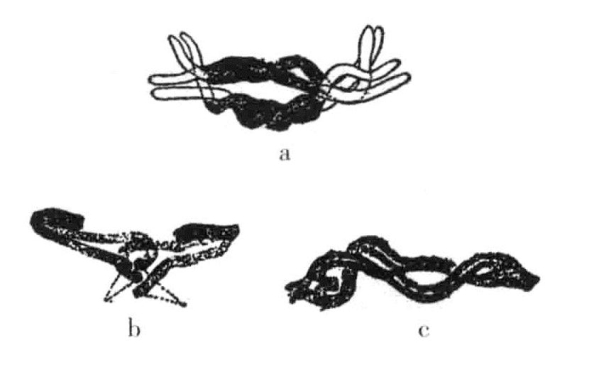
图3-7 Batrachoseps染色体的缠绕
Batrachoseps染色体在进入第一次成熟分裂的纺锤体之前,染色体呈粗丝形状,已是缠绕的晚期(根据Janssens)
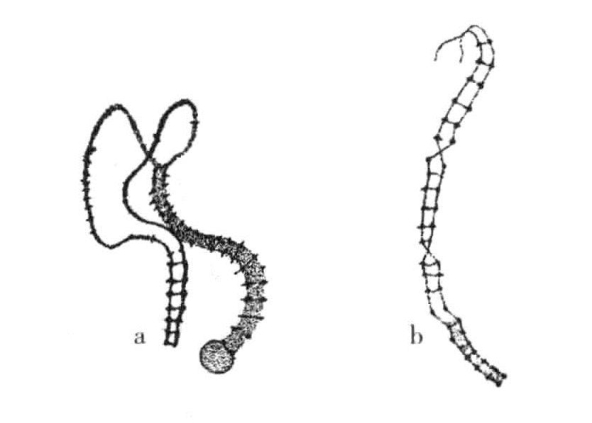
图3-8 涡虫(Dendrocoelum)染色体的接合
涡虫的一对染色体接合。a显示两条细丝正在靠拢;b显示两条接合丝在两个层面上交换(根据Gelei)
最有可能证实细丝缠绕的,是那些开始于一端(或弯曲染色体的两端),并且趋向另一端(或向中间弯曲)的接合。或许以Batrachoseps精细胞这一类最引人注意(图3-6),但是Tomopteris(浮蚕属)图也几乎或完全适用。涡虫卵细胞图(图3-8)也具有充分的说服力。至少在有些图中,给人一种染色体丝向一起靠拢时出现一次或多次重叠的印象,但是这一印象并不足以证明它们除在某种平面上表现出交叉外会产生更多的联系。此外,这并不表明它们重叠的部分一定发生交换。同时必须承认,还没有细胞学证据能证实交换,就这一状况的本质而言,实际上也很难精确地证实。虽然如此,很多例子证明,可以假定进入接合位置的染色体可能很容易发生交换。
因此,细胞学家对染色体的描述,在某种程度上满足了遗传学的要求。回顾这一事实,即在发现孟德尔的论文之前,已经发现大量细胞学证据,并且没有一项工作的进行是带有遗传学偏见的,而是独立于遗传学者的研究的。这些关系似乎不能只是巧合,而更有可能是学者们已经发现了遗传机制的许多重要部分。通过这种机制,遗传要素按照孟德尔两条定律分配,同对染色体有秩序地交换。