近缘物种基因的位置
德弗里斯的突变理论,除了第五章讨论的特殊解释之外,还假设“初级”物种是由大量同质基因构成的,同时认为这些基因按照不同方式重新组合,导致了“初级”物种之间的差异。近来对同属物种杂交的研究,已经为这一理论提供了一些证据。
研究这一问题最容易的方法是让不同的物种杂交,并在可能范围内判断它们是否都是由相同数量的同类型基因构成的。但是,这样做存在一些困难。例如,很多物种是不能杂交的,而且有的物种即便可以杂交,得到的杂交种也有一些是没有繁殖能力的。不过,也存在少部分物种可以杂交,并产生具有繁殖能力的杂交种。即便这样,还存在一个困难,即如何从两个物种中分辨出那些以孟德尔式对的形式出现的性状。每一个例子中的两个物种间存在的各种差异,都受制于很多因子。换言之,在截然不同的两个物种之间,我们很少看到一个差异是起源于一个分化基因的。因此,我们必须通过一个或两个物种中新出现的突变型差异,来为此提供证据。
在若干植物案例与至少两个动物案例中,突变型物种与另一物种杂交,得到具有繁殖能力的子代。这些子代自交或回交,其结果为确定不同物种间的基因等位关系提供了唯一具有决定意义的证据。
East使两种烟草(Nicotiana Langsdorffii与N.alata)杂交(图7-1),其中一株是开白色花的突变型。虽然其杂交孙代的性状变化不小,但有四分之一的植株仍开白花。由此可知,一个物种的突变基因作用于另一个物种的基因,就如同作用于同一个物种的正常等位基因一样。
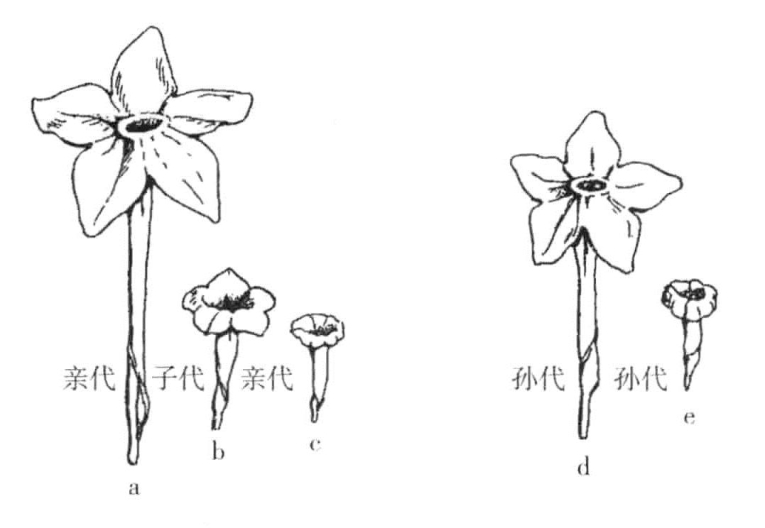
图7-1 两种烟草的杂交
Nicotiana Langsdorffii与N.alata两种烟草的杂交:a、c表示两种原型花;b表示杂交种型花;d和e表示两类孙代回复型花(根据East)
Correns用两种紫茉莉Mirabilis Jalapa与M.longiflora杂交。这里选择了Jalapa(chlorina)的一个隐性突变体,在其孙代中,这一性状大致会在四分之一的植株中再度出现。
Baur用两种金鱼草Antirrhinum majus与A.molle杂交(图7-2)。这里至少采用了A.majus的五种突变型,各个性状都以所预期的数字在孙代中重新出现(图7-3、图7-4)。
Detlefsen用两种豚鼠Cavia porcellus与C.rufescens杂交,雄性杂交种没有繁殖能力。再使雌性杂交种与C.porcellus突变型雄性交配,总计产生7种突变性状。突变性状的遗传方式与C.porcellus中的遗传方式是一样的。这一结果再次表明两个物种含有若干相同的基因点。然而,由于还没有对与porcellus的突变性状类似的突变体进行研究,这些结果并不能充分证明两个物种之间存在相同的突变体。
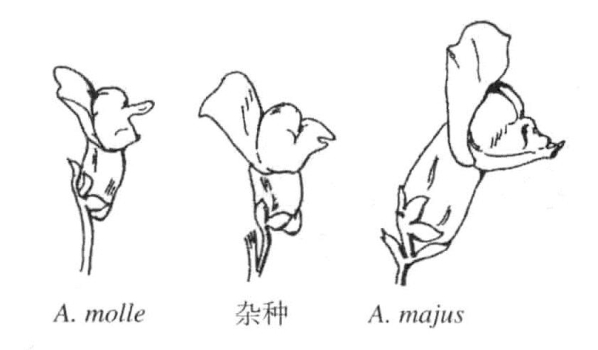
图7-2 两种金鱼草的杂交
Antirrhinum majus与A.molle两种金鱼草及其杂交种(根据Baur)
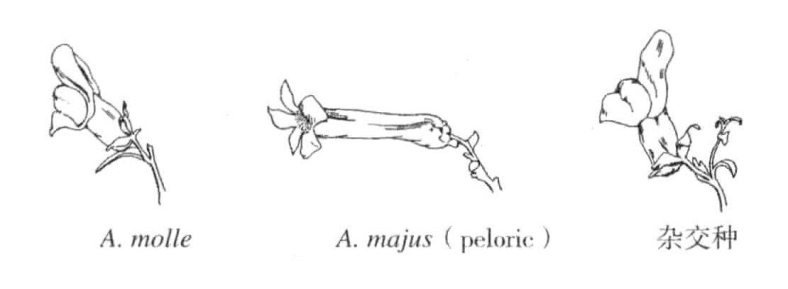
图7-3 突变型金鱼草与正常型金鱼草的杂交
A.molle两侧对称的花朵与A.majus的peloric型杂交,产生下方的野生型杂交种(根据Baur)

图7-4 两种金鱼草(图7-3)杂交产生的不同组合型
图7-3中杂交二代的花朵类型(根据Baur)
Lang在对两种野生蜗牛Helix horte
ensis与H.nemoralis所做的杂交实验(图7-5)中,描述了一个非常明确的例子。这一实验表明,一个物种的性状相对于另一个物种的性状所表现的显性—隐性关系,与同一个物种内同一对性状的显性—隐性关系是一样的。

图7-5 两种蜗牛的变种与杂交
a.Helix nemoralis,00000,黄色,Zurich型;b.同上,00345,带红色,Aarburger型;c.典型H.horte ensis,12345;d.同上;e.杂交种,00000(根据Lang)
有两种果蝇外形极其相似,甚至于曾被认为属于同种。它们是黑腹果蝇(Drosophila melanogaster)和D.simulans果蝇(图7-6)。如果仔细观察的话,我们会发现这两种果蝇之间存在很多不同点。这两种果蝇不容易实现杂交,其产生的杂交种也是完全没有生育能力的。
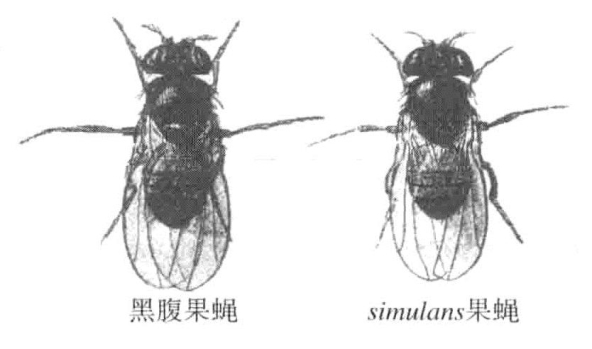
图7-6 黑腹果蝇与simulans果蝇
图左为黑腹果蝇,图右为simulans果蝇,二者皆为雄性
在simulans果蝇中,已知存在42种突变型,分别属于三个连锁群。
simulans果蝇有23种隐性突变基因在杂交种内仍表现为隐性,黑腹果蝇有65种隐性突变基因在杂交种内已被证明是隐性。因此,每一个物种都含有另一个物种中的隐性基因的标准型基因或野生型基因。
在测验过的16种显性基因中,除一种外,其余杂交种内的基因在本物种中似乎都发挥着同样的作用。因此,这一物种的16个正常基因,对另一物种的显性突变基因表现为隐性作用。
对simulans突变型果蝇与黑腹果蝇交配的20个例子所做的检验,已经证明这两个物种的突变性状是一样的。(https://www.daowen.com)
这一结果确定了两个物种在该突变基因上的同一性,人们也可以借此发现这两种突变基因是否位于同一个连锁群内,以及是否在各群内位于一样的相对位置。Sturtevant找出的相同的突变基因点在图7-7中用虚线表示。在第一染色体上表现得十分一致。在第二染色体上只体现了两个相同的基因点。在第三染色体上不是很一致。如果我们假设第三染色体上存在一大段倒置,导致了相应基因点的次序颠倒,则可以解释第三染色体上的不一致现象。
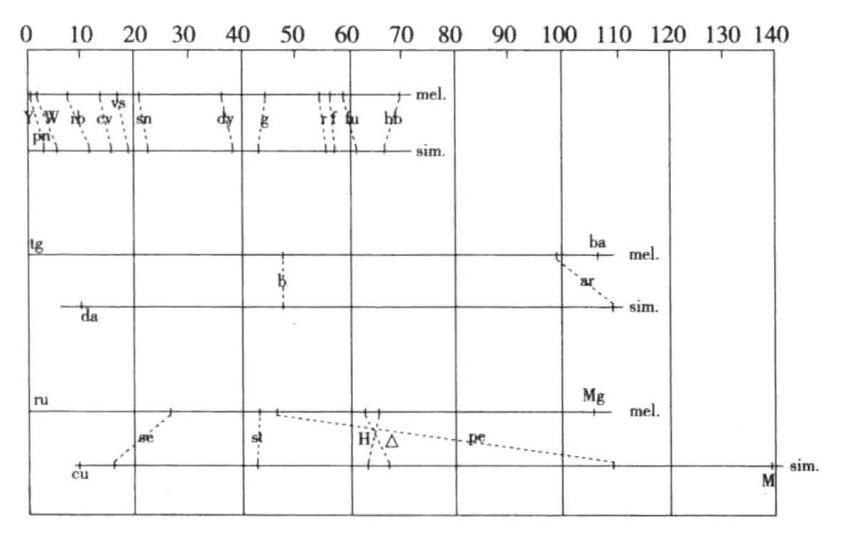
图7-7 黑腹果蝇与simulans果蝇的同型基因
图上部为黑腹果蝇与simulans果蝇第一染色体或X染色体上相同的突变基因对应的基因点,图中部为第二染色体上相同的基因点,图下部为第三染色体上相同的基因点(根据Sturtevant)
Sturtevant的研究不仅本身很重要,也对如下观点有贡献,即认为不同物种间相似的突变基因,只要在连锁群内位于同样的相对位置,都是相同的基因。不过,除非它们经过了像对黑腹果蝇与simulans果蝇所进行的杂交检验一样的验证,否则很难证实其同一性。因为我们已经发现,存在一些相似但不相同的突变类型,并且有的时候它们在同一连锁群内处于相近的位置。[1]
对另外两种果蝇的研究已经发展到一个新的水平,二者间的比较至少是很有意思的。Metz与Weinstein确定了Drosophila virilis果蝇的几个突变基因的位置,Metz把virilis果蝇的基因群的次序与黑腹果蝇进行了比较,图7-8显示virilis果蝇的性染色体上有黄身(y)、无横翅(c)、焦毛(si)、细翅(m)与叉毛(f)这五个明显相似的突变基因,以与黑腹果蝇一样的基因次序进行排列。
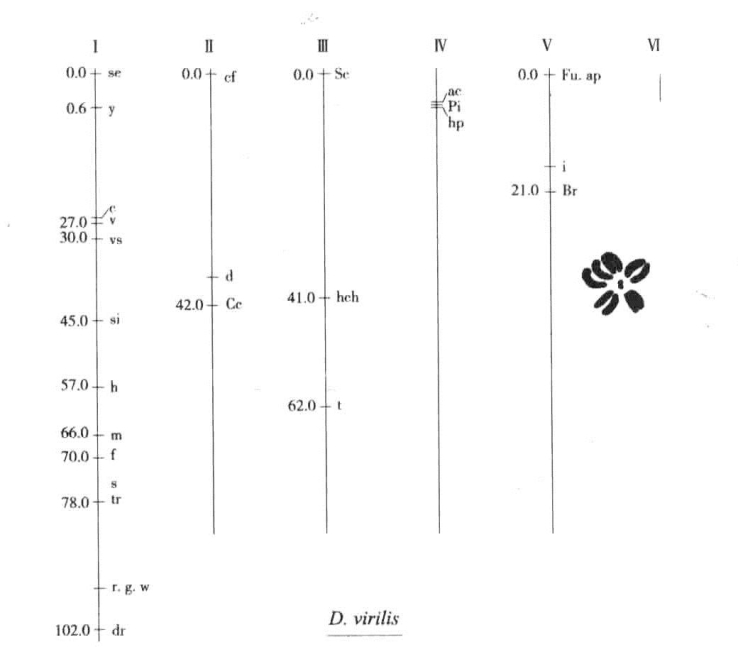
图7-8 virilis果蝇染色体
virilis果蝇6条染色体上突变基因定位表(根据Metz与Weinstein)
据遗传学数据,另一种果蝇Drosophila obscura的性染色体比黑腹果蝇的长一倍(图7-9)。这是很重要的:这条较长的性染色体的中段有黄身、白眼、盾片、缺翅四个突变型基因,与黑腹果蝇较短的性染色体一端的所有同样的突变性状一样。对这一对应关系的解释在Lancefield的严谨的研究中也有体现。

图7-9 obscura果蝇染色体
obscura果蝇染色体上突变基因的位次表,它与黑腹果蝇吻合的基因点为sc=盾片,y=黄身,No2=缺翅,wh=白眼(根据Lancefield)
综合以上各种结果和其他观察结果,我们得出一点认识,即在仅以对染色体群的观察为依据得出系统发育的结论时要严谨。因为通过从果蝇中得出的证据,可以发现亲缘密切的物种,在同一染色体上的基因也可能存在不同的排列次序。相似的染色体群有时也会含有不同组合的基因。既然基因比染色体本身更重要,那么对遗传组成的最终分析,就必然要取决于遗传学,而非细胞学。
[1]考虑到每个基因并不是只发挥一种影响,这些检验工作使基因更有可能被识别出来。