基因的稳定性
以上谈到的,都带有基因是遗传中的一个稳定的要素的意味。至于基因的稳定性究竟是属于化学分子的那种稳定,还是只是在一个固定标准附近定量地变化,却是理论上或根本上的一个重要问题。
既然我们无法借助物理方法或化学方法直接对基因展开研究,我们关于基因稳定性的结论,就只能依据其影响进行推导。
孟德尔遗传理论假定基因是稳定的。它假设各亲体赋予杂交种的基因,在杂交种的新环境中依然是完整的。现在我们通过一些例子来回顾一下这一结论的证据的性质。
Andalusian鸡具有白色、黑色和蓝色三种个体。白鸡与黑鸡交配,得到蓝色鸡。两只蓝色鸡交配,其子代为黑色、蓝色和白色三种,三者具有1∶2∶1的比例关系。蓝色鸡之中,白色基因与黑色基因相分离。一半的成熟生殖细胞得到黑色要素,另一半得到白色要素。任一卵子与任一精子随机受精,经观察,白鸡与黑鸡的孙代的比例关系是1∶2∶1。
可以通过如下测验,来验证杂交种内具有两种生殖细胞的假设是否正确。如果蓝色杂交种回交纯种白鸡,其子代有半数是蓝色鸡,另一半是白色鸡。如果蓝色杂交种回交纯种黑鸡,其子代半数是黑鸡,另一半是蓝色鸡。两次结果都与如下假设相符,即蓝色杂交种的基因是纯净的,一半是黑色基因,另一半是白色基因。二者在一个细胞中,但没有相互影响。
前例中,杂交种与两个亲代不同,从某种意义上说,它属于两个亲代之间的中间型。在第二个例子中,杂交种与一个亲型没有差异。黑豚鼠和白豚鼠交配,其子代都是黑色的。当子代自交时,其孙代的黑白比例为3∶1。孙代白豚鼠与最初的白豚鼠一样,又繁殖出白豚鼠。白色基因虽然与黑色基因在杂交种体内同时存在,却没有受到影响。
还有一个例子,两个亲代很相近,虽然杂交种呈现出某种中间型,但是变化很明显,导致变化的两端分别与两个亲型相重合。在这一类型中,只有一对基因存在差异。
黑檀色果蝇与炱黑色果蝇交配,其子代如上文提到的那样,表现为中间色,并且变化很大。其子代自交,孙代的体色由淡至深,形成一个连续的系列。但是,也有一些办法可以对这些深浅不一的颜色进行检测。检测结果显示,孙代颜色系列是由黑檀色纯种、杂交种和炱黑色纯种组成的,三者具有1∶2∶1的比例关系。由此,我们证明了基因不会混杂的结论。颜色深浅的连续系列只是性状变异性相互掩盖的结果。
上述例子只涉及一对不同的基因,因此显得简单明了。这些例子有助于确立基因的稳定性原则。
然而,实际上,现实情况要复杂得多。很多类型在几个基因上存在差异,每个基因都对同一个性状产生影响。所以,无法从它们的杂交例证中得出简单的比例关系。长穗玉米与短穗玉米进行杂交,子代杂交种的穗轴长短适中。子代自交,孙代出现长短不一的穗轴。有的像最初的短穗族那样短,有的像最初的长穗族那样长。二者属于极端情况。在二者之间存在一系列的中间长度。检验孙代个体,表明有几对基因影响穗轴的长短。
还有一个关于人的体长的例子。人的身体长可能是因为腿长,也可能是因为躯干长,或者两者都长。有的基因可以影响所有部分,有的基因则对某一部分产生更大的影响。结果导致遗传情况复杂。这一问题至今没有得到解决。另外,环境对最终结果也可能存在某种程度的影响。
这些都是多对因子的例子,遗传学者试图弄清楚每个杂交中究竟存在多少因子,结果之所以变得复杂,只是因为所涉及的是几个基因或很多个基因。
在孟德尔的研究被发现之前的一段时期内,这类变异性为自然选择理论提供了依据,我们以后再探讨这一问题。先来看看因为1909年Johannsen卓有成效的研究,我们在认识—选择理论中的一些限制上取得的巨大进展。
Johannsen的实验采用的是一种园艺植物公主菜豆。这种菜豆完全借助自花授粉繁殖。长期连续自交的结果就是每个植株都变成了纯合子,即每对的两个基因都相同。所以,这类素材适合于进行精确的实验,进而确定菜豆的个体差异是否受到选择的影响。如果选择会改变个体的性状,那么在这一情况下,选择必须先改变基因本身才可以。
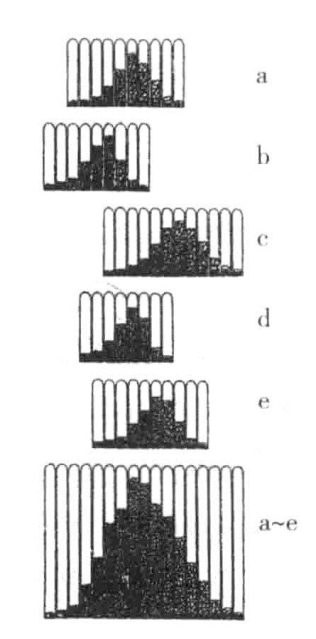
每一植株的豆粒大小都存在些微差异,并且按照豆粒的大小进行排列时,得到正态概率曲线。不管连续从各世代中选出的是大豆粒还是小豆粒,任一植株及其所有后代,在豆粒大小上都表现出同样的分布曲线(图18-1),其后代总是得到同样的一群豆粒。
通过对素材进行检查,他共发现了9族菜豆。他认为其研究成果足以证明某一植株之所以产生大小不同的豆粒,是由于最广义的环境引起的。只要在选择开始时选用每对两个相同基因的素材,就可能证实这一观点。这说明选择对基因本身的改变不会产生影响。
如果一开始选择的有性繁殖的动物或植物不是纯合的,那么直接结果就会不同。这一点在很多实验中都得到了证明,如Cuénot的斑毛鼠实验、McDowell的家兔耳长实验以及East和Hayes的玉米实验。这些实验都可以作为说明在选择下所发生的变化的例证。这里只就其中一个例子进行阐述。
Castle研究了选择对一族披巾鼠毛色的影响(图18-2)。他最初用商品披巾鼠的子代进行实验,一端选择条纹最宽的披巾鼠,另一端选择条纹最窄的披巾鼠,并将两个系列分隔开。经过几个世代之后,这两群披巾鼠表现出一定的差异:一群的背条平均比原来的一群宽,另一群的背条比原来的一群窄。选择作用已经以某种方式改变了条纹的宽度。截至目前,这些实验结果还不能证明这一变化不是选择将决定背条宽度的两组因子分隔开导致的。但是,Castle指出,他研究的是某个基因的影响,因为披巾鼠与全黑色鼠(或全褐色鼠)杂交,子代杂交种再进行自交时,孙代就可以得到全黑色鼠(或全褐色鼠)和披巾鼠两种类型,二者具有3∶1的比例关系。这种孟德尔式比例确实证明了毛上的有色条纹来自一个隐性基因,但未能证明这个基因的作用并非不可能受到与决定背条宽度相关的其他遗传因子的影响,这是真正存在争议的地方。

图18-2 披巾鼠的类型
四种类型的披巾鼠(根据Castle)
之后,Castle还进行了一个由Wright设计的实验,从事实上证明了这些结果是由条纹宽度的修饰基因被分隔开导致的。检验方法如下:经过精选之后的各族,回交野鼠(全身黑色或褐色),得到第二代条纹鼠。再用第二代条纹鼠回交野鼠。经过两三代回交之后,发现被选择的一群,似乎又回到了最初的样子。选择出的窄条纹一族向宽条纹变化,选择出的宽条纹一族则向窄条纹变化。也就是说,精选的两族越来越接近,越来越与其最初的那一族接近。
这一实验结果完全符合如下观点,即野鼠有影响条纹鼠条纹的宽度的修饰基因。也就是说,原来的选择作用,将变宽基因与变窄基因分隔开,进而改变了条纹的属性。
Castle甚至一度声称:披巾鼠实验的结果重建了他所谓的达尔文的观点,认为选择作用本身使得遗传物质沿着选择的方向发生变化。如果这是达尔文的真实意思,那么这种对变异的理解似乎能够大大地巩固进化借助自然选择而进行的理论。1915年,Castle指出:“我们现在拥有的所有证据,都表明外界的修饰因子不能解释在披巾鼠实验中观察到的这些变化,披巾鼠式样本身就是一个明确的孟德尔单元。我们不得不做出如下结论:单元本身在反复选择下,朝着选择的方向变化。有时就像我们的‘突变’族那样突然出现变异,突变族本身是一个高度稳定的正变异,但更多的是逐渐发生变化,就像在正、负两个选择系列中不断发生的那样。”
第二年,他说道:“目前,很多遗传学者认为单元性状是不可以发生改变的……几年来,我研究过这个问题,在这一点上得出一个总的结论,即单元性状可以发生改变,也可以重新组合。很多孟德尔学派的学者持有另一种观点,但我相信,这是因为他们对这个问题的研究还不够精准。单元性状出现定量的变异这一事实是正确无疑的……选择作为进化过程中的一个动力,必须恢复其在达尔文预计中的那种重要性,它是一个能够产生连续进步的种族变化的动力。”
然而,达尔文是否认为选择过程决定或影响了未来变异的方向呢?仔细研读达尔文的论著,除非引用达尔文关于获得性遗传的另一理论,否则并没有任何文字表述能够体现出达尔文持有这一观点。
达尔文坚决崇奉拉马克学说。只要自然选择理论遇到困难,他就会毫不犹豫地运用这一学说。因此,只要愿意,任何人都可以从逻辑上指出(虽然达尔文本人没有将两种学说混为一谈,Castle也没有这样做),当一个有利的类型被选择时,其生殖细胞就像受到了自身产生的泛子的影响,并且能够预计其将按照被选择的那种性状的方向发生变化。因此,每一个新的进展,必将建立在一个新的基础之上。如果围绕这个新的中数(作为一个突破之前限度的中数)出现了彷徨变异,那么在之前的发展方向上,预计会出现进一步的发展。也就是说,自然选择必将按照每次选择的方向发展。
虽然人们可以认为每当达尔文看到自然选择不足以解释事物时,只好援引拉克马学说来为新的进展提供支持,但正如我之前谈到的,达尔文从来没有运用这一论点来支撑自己的选择理论。
不管是自然的还是人工的选择,今天我们都把选择作用看作至多只在原有基因组合能够影响变化的范围内引起变化,或者说选择作用并不能使一群(一个)物种超过那一群原有的极端变异。严格选择可以使一群达到其所有个体都接近原群表现的极端类型。超越了这一点,选择就不能发挥作用了。目前看来,只有依赖一个基因内部发生的新的突变,或者依赖一群旧基因内部的集体变化,才有可能导致进一步的或退一步的永久进化。
这一结论不仅是从基因稳定性理论得来的逻辑推理,也是建立在很多观察基础之上的,这些观察表明当一群生物受到选择时,它们开始变化很快,很快就缓慢下来,不久就停止了变化,并与原群中的少数个体所表现出的某种极端类型相同或相近。
以上只是就杂交种内部的基因沾染以及从选择的观点出发考虑基因的稳定性问题。关于躯体本身可能影响基因组成这一点,只是略微涉及。如果基因受到杂交种躯体性状的影响,那么作为孟德尔第一定律基本假设的杂交种体内基因的精确分离,就一定是不可能的了。
这一结论使我们直面拉马克的获得性遗传理论。我们不准备审视拉马克理论的各种观点,只是想请大家容许我提出需要注意的某些关系。这些关系是在该学说所假设的躯体影响生殖细胞,即一种性状的一个改变能够引起特定基因内部的相应改变的情况下,想要得到的那些关系。我们可以用几个例子来说明其中的主要事实。
黑家兔与白家兔交配,杂交种为黑色,但杂交种的生殖细胞却分为黑色和白色两种,各占一半。杂交种的黑毛不能影响白生殖细胞。不管白色基因在黑色杂交种体内停留多长时间,白色基因总是白色基因。
如果把白色基因看作某种实体,那么假设拉马克理论可以成立,就应该呈现出该基因所寄居的个体躯体性状的一些影响。
可是,如果将白色基因看作黑色基因的缺失,自然就谈不到杂交种的黑毛可以影响到一个并不存在的事物了。对任何信奉存在—缺失理论的人来说,用这一观点来反驳拉马克理论是不够充分的。(https://www.daowen.com)
但是,从另一个角度看也许更合适。白花紫茉莉与红花紫茉莉杂交,产生了开粉色花的中间型杂交种(图1-5)。如果将白色看作某个基因的缺失,那么红色一定起源于某个基因的存在。杂交种的粉色花比红色花颜色淡,如果性状影响基因,那么杂交种体内的红花基因应该被粉色冲淡。这种影响在本例及其他材料中都没有体现。红花基因与白花基因在粉色杂交种内分隔开,没有出现任何躯体影响。
在反驳获得性遗传理论上,另一证据可能更有力。有一种不规则腹缟果蝇,其腹部原本整齐的黑缟或多或少地消失了一部分(图18-3)。从食物丰富、潮湿并带酸性的培养基中最初孵化的果蝇,其腹部黑缟消失得最多。之后,培养基时间越长越干,此时孵出的果蝇就越来越表现出正常的形状,直至最终与野生型果蝇不存在差异。我们在这里遇到了一个对环境影响非常敏感的遗传性状。这一类性状为研究躯体对生殖细胞可能存在的影响,提供了有利机会。
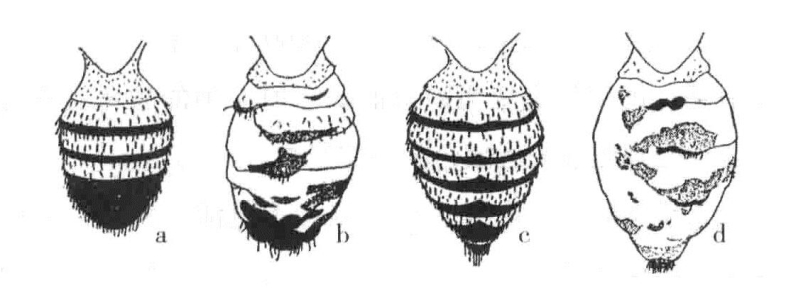
图18-3 果蝇腹部的正常型和突变型
a为正常雄蝇的腹部;b为不规则腹缟雄蝇的腹部;c为正常雌蝇的腹部;d为不规则腹缟雌蝇的腹部
最早孵化出的腹缟高度不整齐的果蝇和较晚孵化出的正常腹缟的果蝇,在相同条件下分别繁育,二者的子代完全相同。最早孵化出的果蝇腹缟不整齐,之后孵化出的果蝇比较正常。对生殖细胞而言,亲代的腹缟是否正常并没有差别。
如果说可能影响太小以至于在子代中观察不到影响,那么补充一句,在较晚孵化出的果蝇连续繁殖了10代之后,仍然看不出存在任何差异。
还有一个确实可信的例证。有一种称为无眼型的突变型果蝇(图4-2)。它们的眼要比正常眼小,并且变化很大。经过选择,得到一个纯粹的原种,其中大多数果蝇无眼,但是,当培养基时间越来越长,有眼果蝇的数量就会越来越多,眼睛也越来越大。如果让这些后孵化出的果蝇繁殖,其子代与无眼果蝇的子代相同。
在不规则腹缟的例子中,较晚孵化出的幼虫,其对称和色素的形成都不是一个明显的存在性状。较晚孵化出的果蝇,其眼的存在是一个正常的性状,这一例子可能被认为比不规则腹缟的例子更合适,但二者的结果是相同的。
近年来,有很多人自以为拿出了获得性遗传的证据,我们没有必要逐一进行验证。我只选出一个有结论所依据的数据和定量的资料,因而也是最完备的例子,即Dürken最近的研究。他的实验应该是仔细推进的,并且对Dürken来说,这个实验提供了一个关于获得性遗传的证据。
Dürken用甘蓝粉蝶的幼虫进行研究。1890年以来,就已确知有些蝶类的幼虫化为蛹(即从幼虫变为静态的蛹)时,蛹的颜色受到背景的一定影响,或者说受到照射光线颜色的一定影响。
例如,如果粉蝶幼虫在白天甚至在弱光下生活、转化,那么其蛹的颜色就会较黑。如果在黄色和红色环境中,或者在黄色和红色的帘后生活、转化,那么其蛹的颜色就会呈绿色。之所以呈绿色,是因为表层黑色素缺失导致内部的绿黄色透过表皮呈现出来(图18-4)。
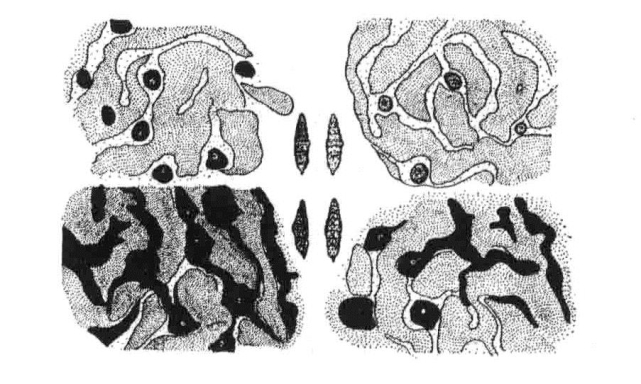
图18-4 菜蝶蛹不同颜色类型的色素细胞分布
图中部显示的是菜蝶4种颜色的蛹。四周表示在不同颜色的类型中,表皮内色素细胞的特殊排列形式(根据Leonore Brecher)
Dürken的实验是在橙色(或红色)光下饲养幼虫,因此得到了浅色或绿色的蛹。将这些蛹转化而成的蝶放置在野外的笼内饲养,并收集它们的卵子。从这些卵子中孵化出的幼虫,有的放在有色光下饲养,有的放在强光或黑暗中饲养。后者作为对照组。实验结果的摘要见图18-5,图中用黑色长实体表示黑色幼虫的数目,用浅色实体表示绿色或浅色幼虫的数目。实际上,蛹可以分成5个有色群,其中3个群合并为黑色群,另外2个群合并为浅色群。
正如图18-5(1)(代表正常颜色)所表示的,几乎所有随机收集的或在自然环境下收集的蛹都是黑色的,只有少数是浅色或绿色的。它们产生的幼虫放在橙色环境中饲养。幼虫转化为蛹,其中浅色类型的百分比很高,见图18-5(2)。如果选出浅色类型饲养,有的放在橙光下,有的放在白光下,其余放在黑暗中,结果如图18-5(3a、3b)所示。图18-5(3a)中的浅色蛹比上一代多,因为在橙光下连续繁殖两代,其影响得以强化。但图18-5(3b)这一组更重要。如图18-5(1)所示,在白光或黑暗中饲养的幼虫比在野生型中出现了更多的浅色蛹。Dürken认为,浅色蛹的增加,一部分在于橙光对上一代的遗传影响,一部分在于新环境的相反方向的影响。
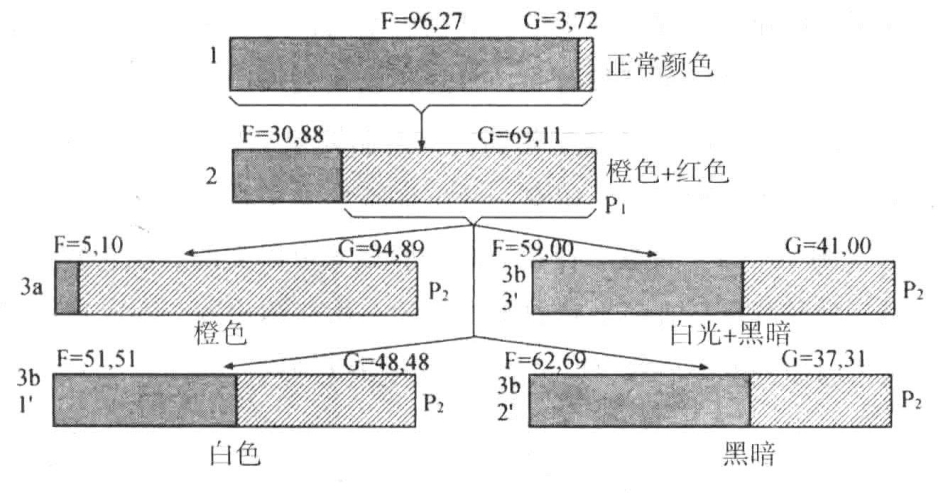
图18-5 在不同颜色的光照下,菜蝶蛹各种颜色类型所占的百分比
图示为菜蝶中黑色蛹和浅色蛹的选择结果(根据Dürken)
从遗传学角度看,这种解释并不能够让人满意。首先,该实验表明并不是所有幼虫都对橙光有反应。如果有反应的幼虫具有不同的遗传性,那么选择它们(实验中的浅色蛹)进行橙光实验,而将对照组放置在白光和黑暗中进行实验时,我们已经在运用一个反应较强的类型,一群经过选择的类型,并且预计它们的下一代将会再次产生反应,而事实也是这样。
所以,除非一开始就采用遗传上同质的素材,或者是采用其他对照实验,否则该证据并不能证明环境的遗传影响作用。
几乎所有此类研究,都犯了同样的错误。即使现代遗传学不能做出更多成果,只是就此类证据毫无价值而言,就可以证明现代遗传学是正确的。
现在我们来看另一些例子,其中有的可能是生殖细胞在经过特殊处理之后受到了直接损害,同时受损害的生殖物质又遗传给后代。由于出现了这一损害,随后各代中可能出现畸形。这意味着这些特殊处理并没有通过首先对胚胎造成缺陷来影响生殖物质,而是同时影响了胚胎及其生殖细胞。
关于酒精对豚鼠的影响,Stockard做了一系列实验。他把豚鼠放置在有酒精的密闭箱中。豚鼠呼吸饱含酒精的空气,几小时之后就完全失去知觉。这个实验进行了很长时间。有的豚鼠在处理中交配,有的在处理结束后交配,二者的结果基本相同。很多胎儿要么流产,要么被吸收,有的胎儿一出生就死亡了,还有一些出现畸形,尤其是神经系统和眼睛的畸形(图18-6)。只有不存在缺陷的豚鼠才可以交配。在它们的子代中,畸形幼龄豚鼠与表面上看起来正常的个体接连出现。在以后的世代中,畸形豚鼠依然会出现,但只能从一定个体中产生。

图18-6 受酒精中毒祖代影响的豚鼠
图示为酒精中毒的祖代产生的两只不正常的幼龄豚鼠(根据Stockard)
对经过酒精处理的谱系进行检查,看不出实验结果符合任何已知的孟德尔式比例的证据。其次,畸形豚鼠呈现出的各部位上的效果,也与单个基因发生改变时表现得不一样。另外,这些缺陷与我们在实验胚胎学中了解到的受到毒物处理后的卵子发育异常很相像。Stockard唤起了人们对这些关系的注意,并认为其实验结果表明酒精使生殖细胞出现某种损害,与遗传机制中某一部分的损害存在关联。影响之所以局限于某些部分,只是因为这些部分对任何脱离正常发育轨道的变化最具敏感性。这些部分以神经系统和感觉器官最为常见。
近来,关于镭射对妊娠鼷鼠和大鼠的影响,Little和Bagg进行了一系列实验。经过适当处理,子宫内的胎儿可能出现畸形发育。产前检查发现,脑脊髓或其他器官(尤其是四肢原基)存在出血区域(图18-7)。这些胚胎有的在产前就已经死亡,并且被吸收了。有的导致流产。有的能够活着出生,其中一部分可以存活并繁殖。它们的后代通常会出现脑或四肢上的缺陷。可能是一只眼或两只眼存在缺陷,也可能没有眼睛或只有一只很小的眼睛。Bagg让这些鼷鼠交配,在它们产生的许多畸形子代个体中,发现了与原来胚胎身上直接引起的缺陷大体类似的缺陷。
图18-7 母鼠遭镭射后,幼鼠的出血区域
母鼠受到镭射后,其子宫内的胎儿呈现出血区域(根据Bagg)
我们应该怎样解释这些实验呢?是不是因为镭射首先影响到正在发育中的胚胎的脑髓,导致缺陷产生,然后由于脑髓的这些缺陷,同一胚胎的生殖细胞才最终受到影响?很显然,这一解释是不成立的。如果只是脑髓受到影响,那么下一代预计会出现脑部的缺陷。如果眼睛是受到影响的主要器官,那么下一代应该只是出现眼部的缺陷。然而在全部报告中,都没有出现此类情况。畸形脑和正常眼的鼷鼠可以产生有缺陷眼的子代。也就是说,这里不存在特定影响,只存在一般影响。
还有一种解释认为,在子宫内时,胚胎的生殖细胞受到镭射的影响。当这些生殖细胞发育成新世代的个体时,这些个体也有缺陷。这是因为最容易在正常发育上受到干扰的器官,就是最容易被发育过程中的任何改变所干扰的器官。总之,它们是最脆弱或者平衡最微妙的发育阶段,因此首先呈现出任何脱离正常轨道的影响。我想,这就是对当前这些实验和其他类似实验的最合理的解释了。