多倍体小麦
小麦、燕麦、黑麦和大麦等壳类植物,都存在一些多倍染色体群。其中,对小麦系的研究最广泛,其杂交后的少数杂交种类型也都经过了检验。单粒小麦的染色体数目最少,为14条(n=7)。单粒小麦属于单粒群,Percival(1921)将其起源追溯到新石器时代的欧洲。还有一种艾美尔小麦群,含有28条染色体,出现在史前时代的欧洲和公元前5400年的古埃及,直到希腊—罗马时代,被含有28条染色体的小麦和软粒小麦群中的一种含有42条染色体的类型(图11-1)所取代。艾美尔小麦群中的变种类型数量最多,但软粒小麦群有着更多的不同类型。
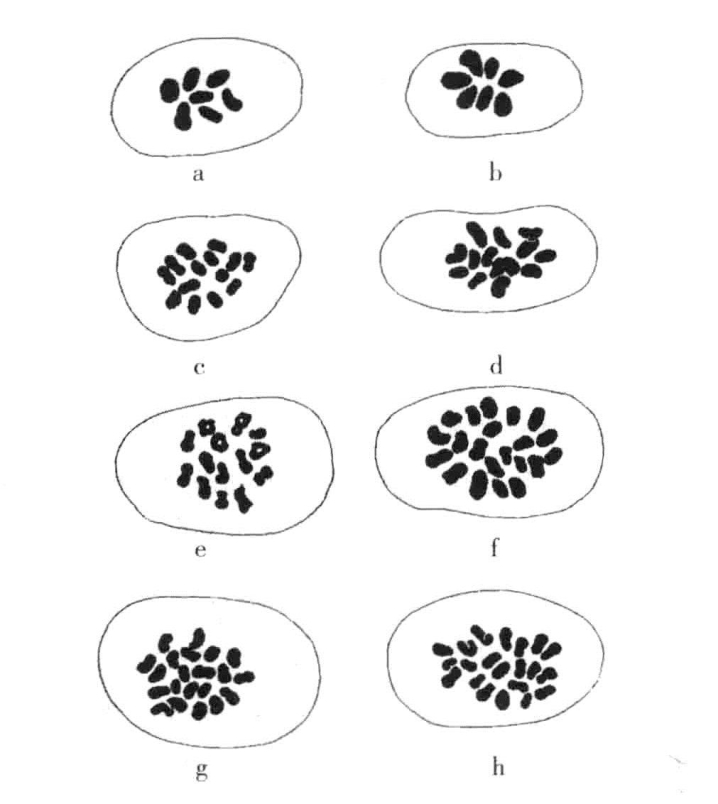
图11-1 二倍体、四倍体、六倍体小麦的染色体
二倍体、四倍体与六倍体小麦减数分裂后的染色体数目(根据Kihara)
有些人对各种小麦的染色体进行了研究,最近的有Sakamura(1920)、Kihara(1919,1924)和Sax(1922)。以下材料大部分来源于Kihara的著作,一部分引自Sax的论文。表11-1显示了观察到的二倍染色体数目与观察到或估计到的单倍染色体数目。
表11-1 二倍染色体数目与单倍染色体数目
各单倍型群参见图11-la(单粒小麦)、图11-1c(坚粒小麦)和图11-1h(软粒小麦)。
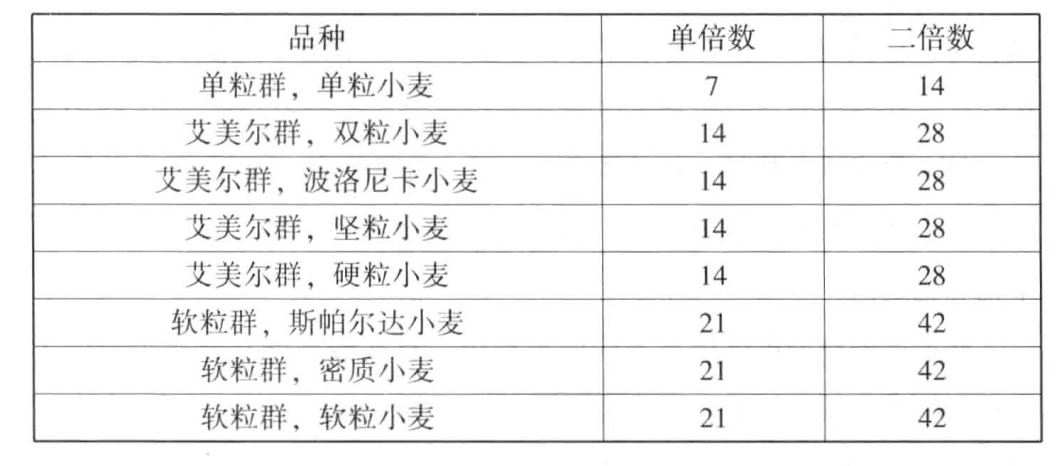
图11-2来自Sax,显示了前述各群的正常成熟分裂。其中单粒小麦有7条二价染色体(接合染色体)。第一次成熟分裂时,二价染色体分裂为二,每一极获得7条染色体,任何一条都没有停留在中途。子细胞进行第二次分裂时,7条染色体分别纵裂为两半,每一极获得7条染色体。艾美尔小麦含有14条二价染色体,第一次成熟分裂时二价染色体各自分裂为二,每一极获得14条染色体。第二次成熟分裂时,染色体分别纵裂,每一极获得14条染色体。软粒小麦vulgare含有21条二价染色体,第一次成熟分裂时,二价染色体分别分裂为二,每一极都获得21条染色体。第二次成熟分裂时,染色体纵裂,每一极获得21条染色体。
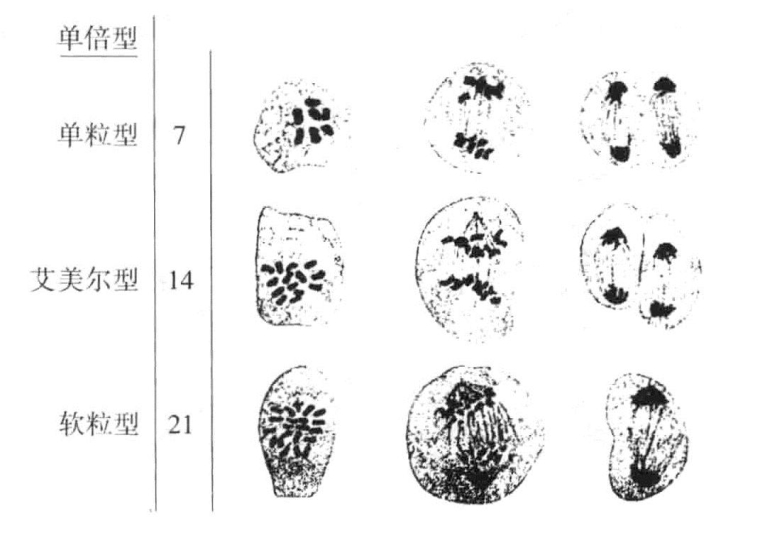
图11-2 单粒小麦、艾美尔小麦和软粒小麦的正常成熟分裂
二倍体、四倍体和六倍体小麦的第一次成熟分裂,即减数分裂(根据Sax)
这一系列类型可以解释为二倍体、四倍体和六倍体。每一类型都是平衡和稳定的。
在上述各种染色体数目不同的类型中,有几种类型在杂交中产生了各种不同组合的杂交种,其中有一些具有一定的生殖能力,有些完全不具备生殖能力。有几种组合的父母双方,各自含有不同数目的染色体。这些染色体的活动,表现出某些重要关系。以下几个例子可以说明这一点。
Kihara研究了艾美尔型和软粒型小麦杂交产生的杂交种。艾美尔型含有28条染色体(n=14),软粒型含有42条染色体(n=21),杂交种则含有35条染色体,因此该杂交种是一个五倍体。当它成熟时(图11-3a~d),含有14条二价染色体和7条单染色体。二价染色体各自分裂,每一极获得14条染色体。单染色体不规则地分布在纺锤体上,并且在“减数”染色体分别进入两极后,依旧停留在原地(图11-3d)。接着,这些单染色体分别纵裂,子染色体趋于两极,但不很整齐。当染色体平均分布时,每一极都能够获得21条染色体。
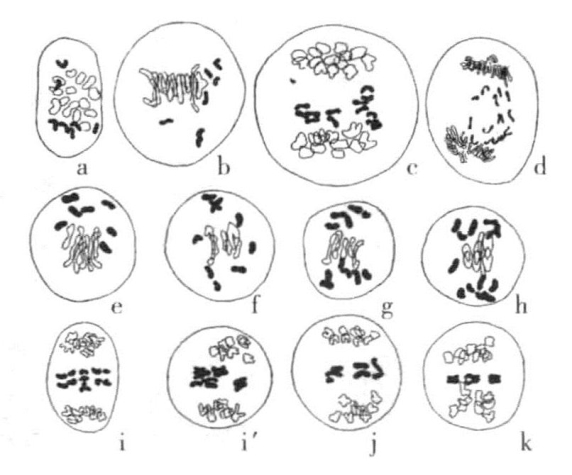
图11-3 艾美尔小麦和软粒小麦杂交种子代的成熟分裂过程
杂交种小麦的减数分裂(根据Kihara)
这里应该提及Sax对三倍体小麦的研究结果,此时,7条单染色体不分裂,而是不均匀地分布到两极,最常见的是3∶4的比例关系(图11-4)。
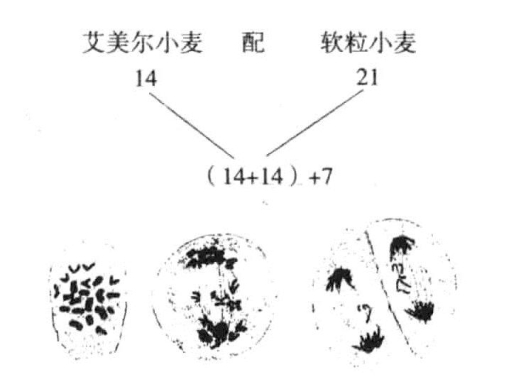
图11-4 同图11-3,对过程的说明稍有不同
图片显示了艾美尔小麦和软粒小麦杂交种的减数分裂(根据Sax)
按照Kihara的研究,第二次分裂时产生了14条正在纵裂中的染色体和7条不分裂的染色体。前者分裂后每一极获得14条染色体,另外7条单染色体呈不规则分布状态,最常见的情况是3条进入一极,4条进入另一极。Sax认为7条单染色体和14条减数分裂后的染色体在第二次分裂时分裂成两条。
不管哪一种解释适用于单染色体(在其他生物中两种解释都有例证),一个明显的重要事实是,只有存在14条染色体时,才能彼此接合。而究竟是艾美尔小麦的14条染色体与软粒小麦的14条染色体接合,还是艾美尔小麦的14条染色体联合成7条接合体,软粒小麦的14条染色体联合成7条接合体而最终都剩下7条单染色体,从细胞学证据上看,这是不能确定的。对这些组合或其他类似组合(可以产生具备生殖能力的杂交种)所做的遗传学研究,也可以提供决定性证据,只是目前尚欠缺此类研究。
Kihara还用含有14条染色体(n=7)的单粒小麦与含有28条染色体(n=14)的艾美尔小麦进行杂交。杂交种是含有21条染色体的三倍体。杂交种生殖细胞(花粉母细胞)成熟时,其染色体的分布比前述例子更为不规则(图11-3e~k)。接合染色体的数目有多有少,染色体之间的接合如果发生的话,也不是完全的。表11-2显示了二价染色体数目上的变化。(https://www.daowen.com)
表11-2 二价染色体数目上的变化
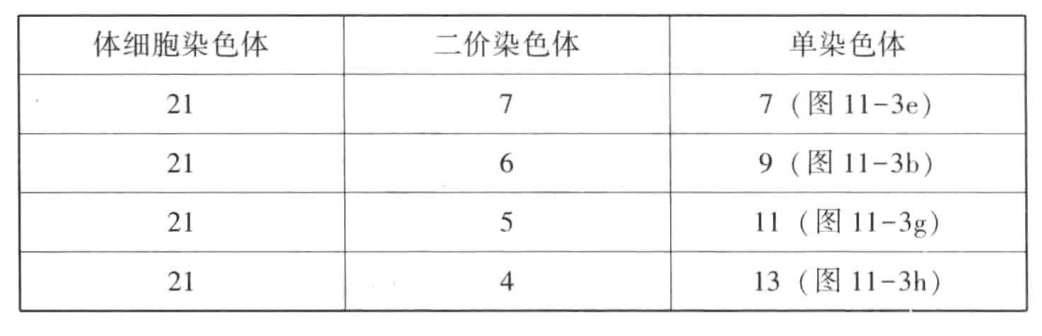
第一次成熟分裂时,二价染色体分裂为2条,分别进入两极。单染色体在没有进入任一极前,不是每一次都会分裂。有的没有发生分裂就进入两极,有的先发生分裂,然后各自进入两极。7条单染色体停留在两极染色体群之间的中央平面上的情况也很常见(图11-3i)。表11-3显示的是三次计量的数字。
表11-3 三次计量的数字
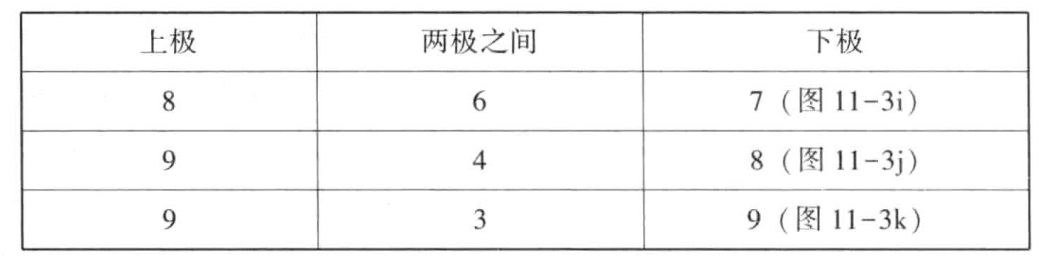
第二次分裂时通常含有11条或12条染色体,有的是二价染色体(纵裂),其余为单染色体。前者正常分裂,子染色体进入一极或另一极。单染色体不分裂就进入其中的一极。
这一证据表明,不能确定杂交种中究竟哪一种染色体接合了。既然二价染色体不超过7条,那么也可以认为它是艾美尔小麦的14条染色体相互接合。
让艾美尔小麦与软粒小麦杂交,可以产生具备生殖能力的杂交种。Kihara对重孙代、玄孙代及以后世代中的一些杂交种成熟分裂时的染色体进行了研究。各植株的染色体数目有多有少,成熟分裂中的一些染色体也呈不规则分布,进而导致更为混乱的情况,或者重新建立起如亲代一样的稳定型。虽然这些结果对研究杂交种遗传而言很重要,但因为过于复杂,与我们的研究目的不吻合。
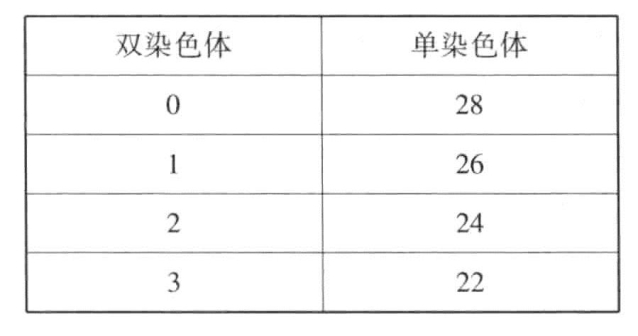
Kihara对软粒小麦与黑麦的杂交种进行了研究,软粒小麦含有42条染色体(n=21),黑麦含有14条染色体(n=7),杂交种(有28条染色体)可以称为四倍体。根据之前的观察,这种由两个不同物种杂交得到的杂交种是不具备生殖能力的,但也有一些观察者认为其具备生殖能力。
生殖细胞成熟时,能够观察到的接合染色体很少,甚至没有,如表11-4所示。
表11-4 生殖细胞成熟时观察到的接合染色体
第一次分裂中,染色体在两极呈不规则分布,只有少数染色体在进入两极之前分裂,有的单染色体散布于细胞质内。第二次成熟分裂时,很多染色体出现纵裂,在第一次分裂时已经分裂的染色体行动缓慢地趋向一极,此时行动缓慢的染色体数目比第一次成熟分裂时行动缓慢的染色体数目少很多。
在小麦与黑麦的杂交中,最令人感兴趣的特点是几乎不存在接合染色体,这会导致染色体的不规则分布,这种不规则分布在很大程度上似乎可以解释杂交种通常表现出的不具备生殖能力的特点。还有一种可能,就是同一物种的所有(或大多数)染色体可能(概率较小)会进入另一极,并产生具备功能的花粉。