回复突变(返祖性)在解释突变过程中的意义
如果说隐性基因的产生是由于基因缺失,那么隐性纯种一定不会再产生原有基因,否则高度特殊的物种居然能够无中生有,这是没有道理的。另一方面,如果突变的发生是由于基因结构的某种变化,那么似乎不难想象有时突变基因能够恢复原态。或许因为对基因了解得过少,我们不能对这一证据的价值给予过高的估计,但是后一种见解对返祖性突变体的出现的解释更合理一些。遗憾的是,这方面的证据还不能令人满意。确实有几个关于果蝇的例子,在突变型隐性原种里产生了具有原有性状或野生性状的个体。但是,除非是在控制之下,否则这种事例还不能作为充分证据,因为我们不能忽视隐性原种受到野生型个体影响的可能性。只有在突变原种中有几个突变性状作为标记,并且只有其中一个性状复原,同时周围没有这几个突变体的其他组合形式的情况下,这种回复变化才会作为令人满意的证据。在培养的原种中,只有少数几个例子符合这些条件。就这些证据所涉及的范围而言,也表明回复现象是可以发生的。同时,我们也要防止另一种可能性的出现,即存在若干突变原种,在经历一定时间之后,似乎或多或少地丧失了原种的特性,可是在杂交之后,又完全恢复了突变性状。例如,第四染色体上的弯翅性状(图4-2),原本是不稳定的,很容易受到外界的影响,如果不经过选择的话,其在外貌上就存在原种野生型的特征。如果此类外貌返祖的果蝇与野生型杂交,其子代再自交,则在所得到的弯翅孙代中,预计会出现很多具有弯翅性状的个体。在另一种盾片突变原种中,也得到了相同的结果。盾片原种具有胸部缺少一些刚毛的特征。在某些盾片原种中就出现了一些缺失刚毛的个体。从表面看,这一突变体似乎是回到了野生型,但当这类果蝇与野生型原种交配时,却证明不是这样的。在杂交产生的孙代中,盾片性状再度出现。通过这一例子,我们发现盾片之所以会还原为正常性状,是因为一个隐性基因的存在,这一隐性基因在盾片原种的纯合状态下,使其缺失的刚毛再度出现。这一结果关系到我们正在讨论的问题。除此之外,一个新的隐性突变促使原有的突变性状回到原型,这也是一个引起人们兴趣的重要事件。
最后,还有一个奇怪的现象,即棒眼可以还原为正常眼。棒眼(图6-7a、图6-7b)是一种显性或半显性性状。很多年来,根据May与Zeleny的观察,我们已经知道棒眼可以还原为正常眼,并且也有人引用这一事例来证明回复突变。回复突变的频率根据不同的原种类型而有所变化,据估算,大约为1/1600。随后,Sturtevant和摩尔根发现,在棒眼回复为正常眼时,棒眼基因的周围就会发生交换。Sturtevant在确定所发生的变化的性质方面得到了决定性的证据。
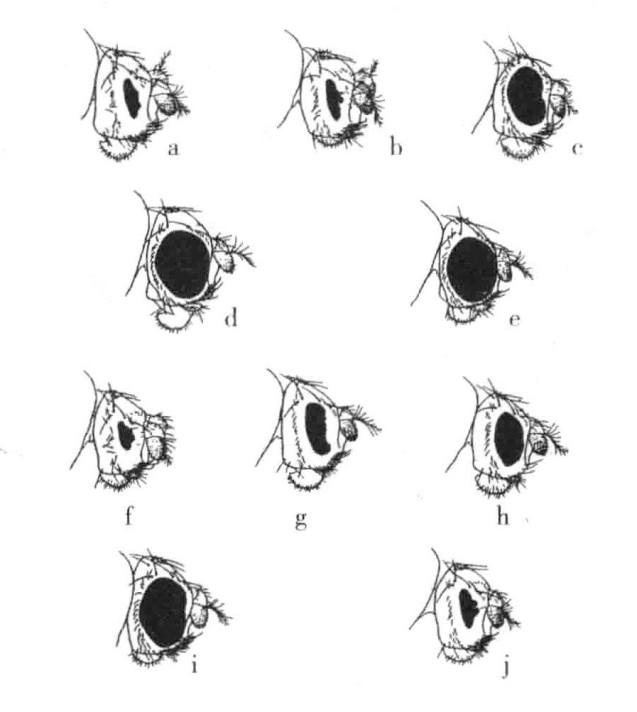
图6-7 果蝇棒眼类型
棒眼的各种类型:a.纯合的棒眼雌蝇;b.棒眼雄蝇;c.棒眼对圆眼的雌蝇;d.回复作用产生的纯合圆眼雌蝇;e.回复作用产生的含有一个圆眼基因的雌蝇;f.双棒眼雄蝇;g.纯合的次棒眼雌蝇;h.次棒眼雄蝇;i.次棒眼对圆眼的雌蝇;j.双次棒眼雌蝇
可以通过以下方法证明每次回复都存在交换,即一个叉毛基因紧贴在棒眼基因的左侧(1/5单位),另一个合脉基因位于棒眼基因的右侧(2.5单位)。一只雌蝇具有如下组合形式,即一条X染色体上含有前述三个基因,棒眼基因位于叉毛与合脉之间;另一条X染色体上不仅含有棒眼基因,还存在叉毛基因和合脉基因的野生型等位基因(图6-8)。这只雌蝇与叉毛、棒眼、合脉的雄蝇交配,普通子代雄蝇从母方接受一条含有叉毛、棒眼、合脉的X染色体,或者接受另一条含有非叉毛、棒眼、非合脉的X染色体,其表现出的性状要么是叉毛、棒眼、合脉,要么只有棒眼。当回复突变发生时(虽然很少发生),即圆眼雌蝇个体出现时,就可以发现叉毛与合脉发生了交换。例如,回复的雄蝇要么具有合脉性状,要么具有叉毛性状,但从未出现过同时具有合脉与叉毛性状或者同时具有非叉毛与非合脉性状的情况。因此,母方染色体上的叉毛与合脉基因之间一定是发生了交换。叉毛与合脉的交换概率不超过3%,但这已经囊括了所有回复突变。
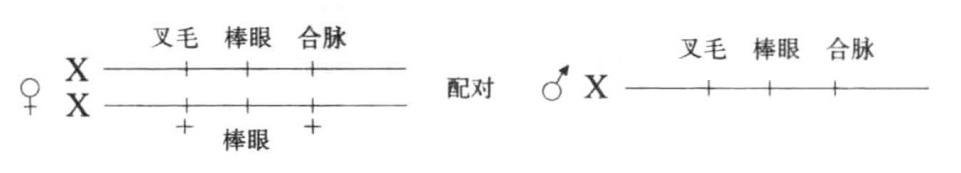
图6-8 棒眼雌蝇在叉毛、合脉上的杂合子与叉毛、棒眼、合脉雄蝇交配
图示棒眼雌蝇在叉毛、合脉上为杂合子,该雌蝇与一只叉毛、棒眼、合脉雄蝇杂交
出于简化情节的考虑,前面只列举了回复型雄蝇的例子。回复型染色体自然也可以进入一个卵子,发育成雌蝇。为了找到回复型雌蝇体内存在交换的证据,我们可以设计一个这样的实验。普通子代雌蝇都是纯合的棒眼(图6-7a),回复型子代雌蝇是杂合的棒眼,要么加上叉毛,要么加上合脉,既没有同时具备叉毛和合脉的个体,也没有同时具备非叉毛和非合脉的个体。
导致回复为圆眼的交换,一定不只是使一条X染色体失去一个棒眼基因,而是一定会将这一基因移放到另一条棒眼染色体上(图6-9a)。含有两个棒眼基因(双棒眼)的雄蝇,与含有一个棒眼基因的雄蝇,在外观上是比较相似的,只是前者的眼睛相对较小,其小眼的数目也相对较少,这种类型我们称其为“双棒眼”(图6-7b)。在同一个直线序列上存在着两个等位基因,这一不同寻常的情况在其他突变情形中从未出现过。可以把这一情况解释为,我们只能假设在交换发生之前,原本对立的两个棒眼基因,在交换发生时略微移动了位置,结果导致双棒眼染色体上至少延长了一个棒眼基因,而另一条染色体则因为缺失一个棒眼基因而相应地缩短了。
Sturtevant用一些具有决定性影响的定性测验来验证回复理论。由棒眼突变得来的一个棒眼等位基因,我们称之为“次棒眼”(图6-7g、图6-7h)。这种类型的果蝇在眼睛的大小和小眼的数目方面都与棒眼型有些微差异。回复现象也会在次棒眼原种中出现(图6-9b),并产生出类似于野生型的完全圆眼型和被命名为“双次棒眼”的新型个体(图6-7j)。
雌蝇在一条染色体上含有棒眼基因,在另一条染色体上含有次棒眼基因(图6-9c)。当回复发生时,该雌蝇会产生完全圆眼型和棒眼—次棒眼型或次棒眼—棒眼型果蝇(图6-9c)。
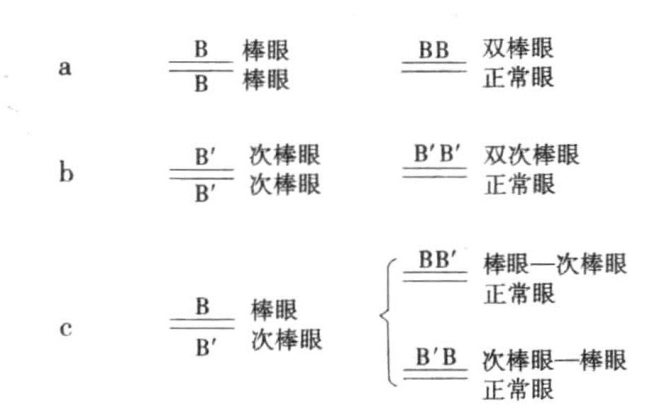
图6-9 棒眼、次棒眼、棒眼—次棒眼的交换
图示棒眼、次棒眼与棒眼—次棒眼三者的突变
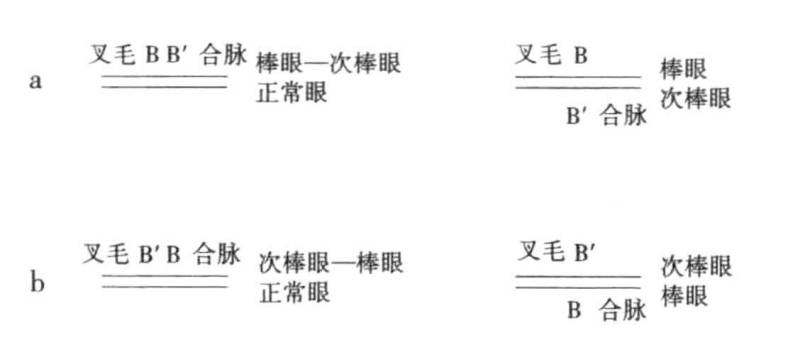
图6-10 叉毛、合脉上的棒眼—次棒眼杂合子与叉毛、合脉上的次棒眼—棒眼杂合子间的交换
a显示叉毛—棒眼与次棒眼—合脉之间的突变;b显示叉毛—次棒眼与棒眼—合脉之间的突变
Sturtevant也通过棒眼—次棒眼型和次棒眼—棒眼型,证明了如下事实:如果突变基因全部位于同一条染色体上,那么当棒眼—次棒眼型与正常型发生交换时(图6-10a),结果产生叉毛—棒眼型或次棒眼—合脉型;如果在次棒眼—棒眼型与正常型之间发生交换(图6-10b),那么结果要么产生是叉毛—次棒眼型,要么是产生棒眼—合脉型。
因此,我们得知这两种类型中的各个基因不仅保留了各自的特征,也会维持基因之间的顺序。从f BB'fu和fB'Bfu(f为叉毛基因,fu为合脉基因。——编者注)的构成方式,我们可以了解各基因间的顺序。实际上,在全部例子中,B与B'的分离都与已经确定了的基因顺序相吻合。
这些结果为棒眼是因为交换而得以回复这一说法提供了有力的决定性证据。目前,这是唯一的案例。似乎在X染色体上的棒眼基因点上存在某种特殊情况,促使了等位基因之间发生交换,Sturtevant将其称为“不等交换”。[1]
前述结果还提出了一个问题,即是否所有的突变都是因为交换才出现的。果蝇的例子显然证明了交换不能作为所有突变产生的普遍性解释,因为众所周知,雄蝇是没有交换的,而突变在雌雄果蝇身上都能够发生。