隐性性状与基因缺失
毫无疑问的是,在“存在—缺失”理论之下,隐含着这样一种观点,即很多隐性性状都是原型固有的某些性状的真实损失;从而又推论出各性状的基因也缺失了。这一观点是魏斯曼关于定子与性状的关系理论的残存理念。
对那些看起来似乎能支持该观点的证据做进一步检验,是很有必要的。
人们可以把白兔、大白鼠或白豚鼠说成是失去了原型所特有的色素。在某种意义上,我们不能否认二者之间的关系,这种表述也没有问题。不过需要注意的是,在很多白豚鼠的脚上或脚趾上,还保留有少许有色毛。如果产生色素的基因真的不存在,而毛的颜色又取决于该基因,那么就难以解释有色毛的存在了。
有一种果蝇突变族只有翅的痕迹,称为痕迹翅(图1-10)。但如果该型幼虫在31℃左右的环境中发育,则两翅就会变长,最长的能达到几乎与野生型翅长相同的程度。如果决定长翅的基因真的不存在,那么高温环境如何促使其重新生长呢?
还有一族精选果蝇,大多数都是无眼,少数是小眼(图4-2)。培养的时间越长,有眼果蝇越多,眼睛的平均体积也越大。基因不可能随着培养时间的延长而发生变化,如果最初产生的无眼果蝇缺失该基因,那么长时间的培养也不会恢复缺失的基因。即便我们假设可以恢复缺失的基因,那么培养时间长的果蝇应该产生更多有眼或更大眼的后代。但是,事实上并没有这样的事情发生。
还有其他隐性突变型,其性状缺失本身并不明显。黑兔相对于灰色野兔而言,黑色属于隐性性状。事实上,黑兔比灰兔拥有更多色素。
存在产生纯白个体的显性基因,如白色来亨鸡就是这类因子导致的。这里存在一种相反的观点,据说野生型原鸡有一种抑制白色羽毛的基因,失去该抑制基因,就能产生白色羽毛。这种说法看起来能够自圆其说,不过其假设原鸡含有此类基因的说法还是比较牵强的。同时,如果从其他显性性状的角度看,这一观点是站不住脚的,只能算作为了保住该理论而牵强附会的臆想而已。
我们应该记得,隐性基因与显性基因的差异大多都是勉强划定的。经验表明,性状并不一直是隐性或显性的,相反,在很多例子中,一种性状既不能完全归为显性,也不能完全归为隐性。换一种说法,含有一种显性性状与一种隐性性状的杂交种,大致上是介于二者之间的,即两种基因对所产生的性状都会具有一定影响。如果认清这一关系,那么认为隐性基因是一种缺失的理论就有问题了。当然,在这些例子中,也许有一些理由认为杂交种之所以具备中间性,是因为一个显性基因的影响比两个显性基因的弱,但这一解释又引入了一个新的因素,也不能说这一影响真的起源于某种缺失。我们或许可以迎合这一假设,不过这样做是不必要的。
如果我们承认上述论证成立,那就不必从字面意义上解释隐性基因的含义。不过,近年来出现了另一种对全部基因的影响与性状之间关系的解释。这一解释使反驳存在—缺失理论变得更困难。例如,假设染色体真的失去了一个基因,同时假设当两条此类染色体结合时,个体的某一性状就会改变,甚至缺失。这一改变或缺失,或许可以说是全部其他基因联合产生的影响。促成这一结果的并不是基因缺失本身,而是当某两个基因缺失时,其他基因所施加的影响。这一解释避免了那种相当幼稚的假设,即每一基因都单独代表个体的一个性状。
在探讨这一观点之前,应该指出的是,在某些方面,这一观点与我们所熟知的关于基因与性状之间关系的另一种解释类似,实际上,这一观点是从后一种解释演化而来的。比如,如果我们把突变作用理解为基因组织内部的一种变化,那么当两种隐性突变基因存在时,新性状并不仅仅起源于新基因,而是由包括新基因在内的所有基因共同作用的最终结果,这与原有性状起源于发生突变的原有基因与其他基因的解释是相同的。
简单地说,上述两种解释的意思是:第一种认为,当一对基因缺失时,所有其他基因都会出现突变性状;第二种认为,当一个基因的组织出现变化时,新基因与其他基因共同产生的最终结果,才是突变性状。
最近得到的一些证据,虽然不能对上述两种解释提供决定性的依据,但对解决争论中的一些问题也是有一些作用的。因为这些证据揭示了某些没有讨论过的突变的可能性,所以探讨这些证据本身是有必要的。
果蝇中存在几个突变型原种,它的翅端有一个或多个缺口,其第三翅脉更粗(图6-2),所以统称为缺翅型。拥有此种性状时,只有雌蝇能够存活,凡是携带缺翅基因的雄蝇都没有存活。缺翅基因位于X染色体上。缺翅雌蝇的一条X染色体上存在缺翅基因,另一条X染色体则携带正常的等位基因(图6-3)。缺翅雌蝇的成熟卵子中,有一半含有一条X染色体,另一半则含有另一条X染色体。当这种类型的雌蝇与正常雄蝇受精时,含有X染色体的精子与含有正常X染色体的卵子结合,繁殖出正常雌蝇;含有X染色体的精子与含有缺翅X染色体的卵子结合,繁殖出缺翅雌蝇;含有Y染色体的精子与含有正常X染色体的卵子结合,繁殖出正常雄蝇;含有Y染色体的精子与含有缺翅X染色体的卵子结合,其后代不能存活。最终结果是:杂交子代的果蝇雌雄比例为2∶1。
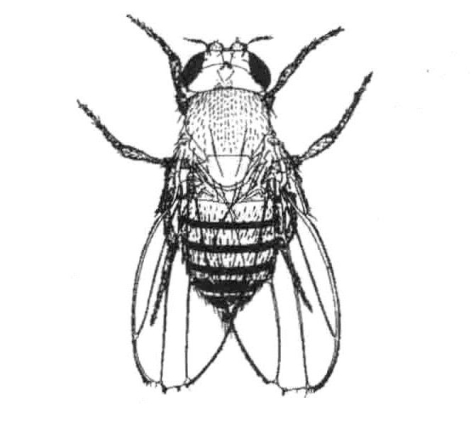
图6-2 缺翅突变型果蝇
黑腹果蝇的缺翅是一个显性的性连锁性状,也是隐性致死性状
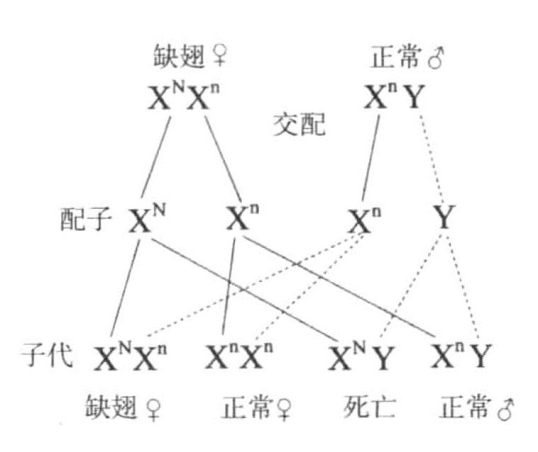
图6-3 缺翅的遗传
图示缺翅雌蝇XNXn与正常雄蝇XnY的杂交,XN代表缺翅的X染色体,Xn代表另一条含正常等位基因的X染色体
单就这一证据来说,或许可以认为缺翅基因是一种隐性致死基因,在杂交种体内,起到了显性翅形修饰基因的作用。但是,在之后的研究成果中,Metz和Bridges(1917)以及Mohr(1923)先后证明了,X染色体上的缺翅突变涉及的范围,要多于普通“基点突变”影响到的部分。因为如果在一条X染色体上相当于缺翅的部分存在隐性基因,而在另一条X染色体上有缺翅基因,那么这个个体就会表现出隐性性状,就如同缺翅染色体上的某一段已经缺失或至少不能存活似的(图6-4a)。事实上,结果就如同真的发生了缺失一样。在某些缺翅突变体中,损失的部分长约3.8个单位(从白眼基因左侧到不规则腹缟基因右侧,如图1-19所示)。在另一些缺翅个体中,损失的部分单位较少。在任何一种情况下,测试结果似乎都表明在某种意义上,染色体或多或少都已经失去了一小段。
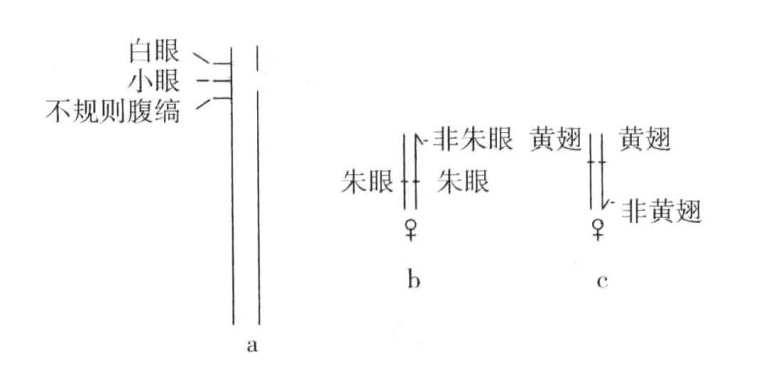
图6-4 作为缺陷的缺翅,非朱眼的复制,非黄翅的复制
a显示缺翅染色体上基因的位置:图右所示的染色体的断裂处为缺翅,图左所示为一条染色体上白眼、小眼与不规则腹缟三种隐性基因与缺翅相对的位置;b显示一段染色体易位到另一条染色体上:两条X染色体各含有朱眼基因,其中一条与朱眼的正常等位基因,即非朱眼基因的一段相连接;c显示两条含有黄翅基因的X染色体,其中一条与黄翅的正常等位基因,即非黄翅的一段相连
正如前面提到的,一些隐性基因与缺翅基因相对时,就会出现它们的隐性性状。不管是把这些隐性基因看成缺失,而由所有其他基因产生影响,还是认为这些隐性基因与所有其他基因一起产生影响,两种说法都与事实相符。实验结果不能判定二者谁对谁错。(https://www.daowen.com)
但是,这一区域中的两个隐性基因产生的性状,与一个隐性基因和缺翅基因缺失所产生的性状之间存在着微小差异。这一差异的存在,或许是由于一种缺失基因(缺翅)加上一个隐性基因,并不等同于两个隐性基因。但是,在进一步研究之后证明,在缺失的缺翅一段中,缺少了某些基因,而在隐性基因中,却含有这些基因,二者的差异可能是由这些基因的存在或缺失导致的。
在上述例子中,只是根据遗传学证据,推导出缺翅突变体的X染色体少了一段,还不能从细胞学方面予以证明。下面的例子则证明了一个真正的缺失。
有时果蝇会缺失一条第四染色体(单倍-Ⅳ,图4-1),一些突变原种的第四染色体上存在几个隐性基因。我们可以造出一类个体,在它唯一的第四染色体上只有一个诸如无眼的隐性基因。这类个体表现出无眼原种的特征,不过作为一种类型,要比存在两个无眼基因的情况更极端。可以认为这一差异是由于失去的一条染色体上缺失了其他基因导致的。
在Bridges与摩尔根(1923)命名为“易位”的例子中,出现了另外一种关系。易位(根据遗传学证据)指的是染色体的一段脱离了该染色体,重新连接在另一条染色体上。该段染色体继续存在。由于其带有一些基因,所以增加了遗传结果的复杂程度。例如,正常染色体上相当于朱眼基因点的一段,易位连接到另一条X染色体上(图6-4)。一只雌蝇的两条X染色体上各含有一个朱眼基因,其中一条X染色体与易位的一段染色体相连接。这样的雌蝇在那一段染色体上具有了朱眼的正常等位基因,但其眼睛的颜色仍旧是朱色。如果我们一开始把朱眼基因当作缺失来看待,那么两个缺失相对于一个存在,应该不会成为显性。但是,我们进一步研究之后,可能会得出另一种解释:如果朱眼是由朱眼基因缺失时所有其他基因的联合作用导致的,那么即使存在一个显性的正常等位基因,也会出现同样的结果。这种情况与一条染色体上含有一个朱眼基因,另一条染色体上含有朱眼基因的正常等位基因的情况不能等同。
这里提到的两个隐性基因与移接段上的一个显性基因的关系,不是总能引起隐性性状的发育。例如,L.V.摩尔根报告的另一个易位的例子,X染色体上相当于黄翅和盾片这两个突变基因所在区域的一段,移接到另一条X染色体的右侧。如果一只雌蝇的两条染色体上分别含有黄翅与盾片基因,其中一条X染色体与易位的一段相连接,那么该雌蝇就会显现出野生型性状。在这一例子中,移接段上的显性等位基因抵消了两个隐性基因的影响。这意味着,所有其他基因与移接段上的各种基因,联合改变了对显性型发育不利的态势。从以上任一学说的角度出发,都可以出现这种现象。
也有对玉米的三倍体胚乳与一种三倍型动物的研究,探讨两个隐性基因与一个显性基因的关系。玉米种子胚乳的细胞核由一个花粉核(含有单倍染色体)与两个胚囊核(分别为单倍)共同构成,最终得到一个三倍型核(图6-5),随后经过分裂,形成了胚乳细胞的三倍型核。粉质玉米胚乳由柔软轻盈的淀粉组成,石质玉米胚乳由大量角质淀粉组成。如果用粉质玉米为母方(胚珠),石质玉米为父方(花粉),那么其子代种子都含有粉质胚乳。因此,得出的结论是,两个粉质基因对一个石质基因,表现出显性作用(图6-6a)。如果按照相反方式交配,以石质玉米为母方,粉质玉米为父方,二者的杂交子代种子的胚乳都是石质的(图6-6b)。在这一例子中,两个石质基因对一个粉质基因,也表现出显性作用。这两种基因中,究竟把哪一种当作另一种的缺失,实际上是任意选择的。如果缺失的是粉质基因,那么在第一个例子中,两个缺失对一个存在而言就是显性的。在第二个例子中,两个存在对一个缺失而言也是显性的。
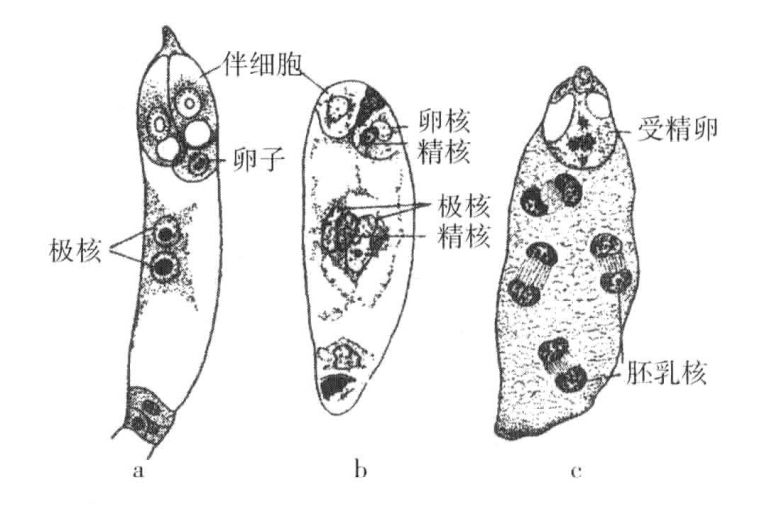
图6-5 显花植物卵细胞的受精,胚乳的发育
a显示植物胚囊内部卵核受精的三个阶段;b显示母方两个单倍型核与父方一个单倍型精核;三核结合在一起,产生三倍型胚乳,如c所示(根据Strasburger与Guinard,见Wilson)
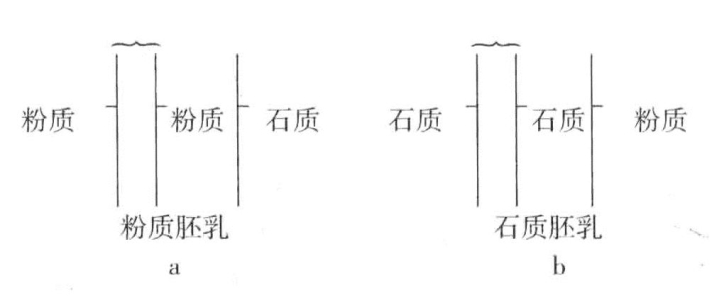
图6-6 三倍型:a为两个粉质基因与一个石质基因;b为两个石质基因与一个粉质基因
图示玉米胚乳的三倍型。a显示两个粉质基因与一个石质基因结合,产生粉质胚乳;b显示两个石质基因与一个粉质基因结合,产生石质胚乳
要是只从字面意思来解释两个缺失基因比一个存在基因占优势,这种说法没有什么意义。不过,就像前面所说的,一个基因缺失时其他基因联合决定粉质性状的说法似乎还有些合理性。或者假设粉质基因存在(由石质基因突变而来),该基因与其他基因一起产生影响,这种说法也行得通。因此,从三倍型胚乳得到的证据,并不能决定隐性究竟是源自于某一基因的缺失,还是存在于某基因。这就如同一段染色体移接时会增加第三者的易位一样。
有一些关于玉米的例子,其中两个隐性要素对一个显性要素,并不占有优势。只是这些例子与当前问题无关。
如果三倍型雌蝇的两条X染色体上各有一个朱眼基因,另一条X染色体上有一个红眼基因,那么将得到红眼性状。在这一例子中,一个显性基因对两个隐性基因占有优势。这一结果与重复段上的一个野生型基因对两个朱眼基因的结果相反。但是,三倍型雌蝇几乎增加了一整条X染色体,重复型果蝇则只增加了一小段X染色体,因此二者存在差异。多出的X染色体上的基因过剩,这可以解释二者之间的差异。隐性基因不管是解释为缺失,还是解释为突变基因,都是可以说通的。