环境变化
Giard在1886年证明了如果雄蟹体上寄生了其他壳类如Peltogaster或蟹奴,雄蟹的外部性状就会发育成雌性类型。图17-la显示了雄蟹成体及其大型螯足;a'表示从腹面观察到的腹部和交配附肢;b表示雌性成虫,螯足较小;b'表示从腹面观察到的腹部和多刚毛的二叶状抱卵附肢;c表示早期受到感染的雄蟹,螯足小,类似雌性,腹部宽,与雌性相同;c'表示从腹面观察到的受感染的雄蟹的腹部,具有与雌性相同的小型二叶状附肢。
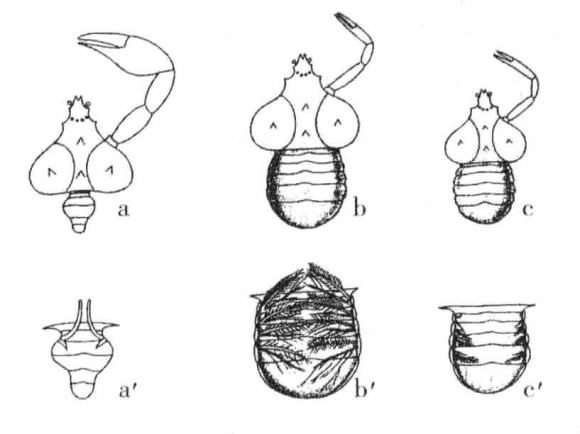
图17-1 蟹的正常雄性、正常雌性和被寄生雄性
蛛蟹:a为正常雄蟹;a'为正常雄蟹的腹视图;b为正常雌蟹;b'为正常雌蟹的腹视图;c为被寄生的雄蟹;c'为被寄生的雄蟹的腹视图(根据Geoffrey Smith)
寄生虫长长的根状突起侵入蟹体内部,以吸食蟹的体液为生,同时能够反过来影响蟹本身的生理过程。蟹的精巢最初可能不受影响,但后来就逐渐退化了。Geoffrey Smith至少在一个寄生虫脱离蟹体的例子中,发现再生的精巢内出现了大生殖细胞。他认为那是卵子。
Giard没有就蟹体变化究竟是因为精巢被吸收还是因为对宿主产生了更直接的影响这一问题得出结论。Geoffrey Smith提供了一些有关血液中脂肪的证据,并拿出一定的证据来支持蟹体发生变化是因为蟹体受到了生理影响这一观点。目前还没有关于甲壳类生殖腺的破坏是否影响第二性征的证据。
在摘除一些昆虫的生殖腺方面已经有了证据,证明摘除精巢或卵巢,不会改变第二性征。因此,Kornhauser(1919)描述的一种甲虫(Thelia)的例子显得更有意义。Thelia被一种膜翅类(Aphelopus)寄生时,雄虫表现雌性的第二性征,至少不能产生雄性的第二性征。
甲壳类的十足目生物大多是雌雄异体的,但也存在少数几个例子,在一种性别或两种性别中,卵巢和精巢同时存在。还有少数几个例子,在幼龄雄虫的精巢内可以观察到大型卵状细胞。也有几个关于蝲蛄的性中型的研究,但还不清楚其是否存在完全的逆转。[1](https://www.daowen.com)
有几位观察者(Kuttner、Agar和Banta等)对水蚤及与其亲缘关系较近的类型中的性中型进行了描述,但没有看到完全逆转的例证。Sexton和赫胥黎(1921)最近对一些被称作雌性性中型的水虱个体进行了描述,它们“在成熟时,多少会与雌性相近,但会逐渐地越来越与雄性相近”。
大多数的藤壶都是雌雄同体的。但在一些属中,除了固定的大型雌雄同体类型外,还有一些微小的起辅助作用的雄性,还有少部分物种只有固定的雌性和起辅助作用的雄性。通常将固定的个体看成真正的雌性,但Geoffrey Smith认为自由游动的幼虫固定下来之后,就会生长壮大,经过雄性阶段,然后变成雌性。但是,如果自由游动的幼虫附着在雌性个体上,就只发育到雄性阶段为止。这似乎只能表明一个未发育的个体发育成雌性,还是终止发育而成为雄性,是取决于环境的。
最后一个例子与Baltzer描述的后螠Bonellia的例子很相近。自由游动的后螠幼虫如果附着在雌螠的吻上,就会非常细小,产生精巢。但如果幼虫单独定居下来,就会发育成一个大型的雌性个体。这个证据并不排斥两种个体各自向着一个方向分化的可能性,但Baltzer的解释看起来是很有可能的。
如果上述解释就是对藤壶和后螠的正确解释,那么这就意味着这些类型的性别是由环境条件决定的,从基因方面看,这表明所有个体都是一样的。[2]