第十章 单倍体
遗传学证据指出,要至少具有一整组染色体,才能展开正常的发育过程。含有一组染色体的细胞称为“单倍型”细胞。由这类细胞构成的个体有时称为“单倍体”,或者称为“单倍型”。胚胎学证据指出,一组染色体是发育的必要条件。但是,并不能因此推断,就所涉及的发育条件而言,在不产生严重后果的情况下,单倍染色体能够直接取代二倍染色体。
经过人工刺激之后,卵子可以发育成胚胎,这类胚胎细胞只含有一组染色体。由于卵子在发育前,胞质的分裂受到抑制,导致染色体增加到两倍的例子也比较多,同时其状态比单倍体好。
切一片海胆卵子,让其与精子受精,可以得到一个含有父方单组染色体的胚胎。Spemann与后来的Baltzer,两人在蝾螈卵子受精后,立刻中缢卵子,有时会分离出一片只含有一个精核的卵子胞质(图10-1)。Baltzer将其中一个培育到变态时期。
用X射线或镭射线照射蛙卵足够长时间,损伤或破坏其染色体,随后让其受精,正如Oscar Hertwig和Gunther Hertwig所证明的那样,这些受精卵也可以发育成由含有半组染色体的细胞构成的胚胎。相应地,蛙的精子在经过射线照射后,虽然仍可以进入卵子,但不再对发育起作用。在这种情况下,卵子只含有卵核的单组染色体,染色体能够分裂,但胞质不能分裂,进而在发育开始之前细胞就恢复了正常染色体的数目。这些卵子在经历了胚胎时期后,能够发育成正常的蝌蚪。
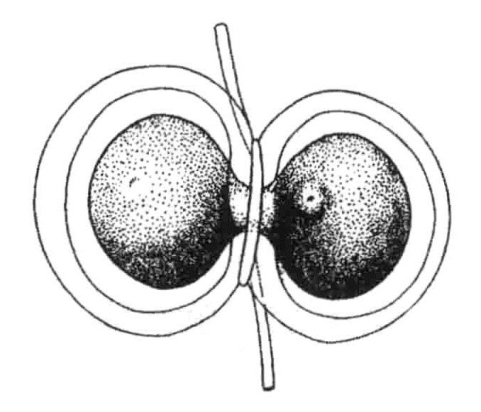
图10-1 蝾螈卵子中缢后,半数与一个精子结合
蝾螈卵子在受精之后,立即中缢为二,右侧的一半表示极体(根据Spemann)
通过上述方法培育的人工单倍体,多数比较衰弱,很多在成年之前夭折。造成这一局面的原因尚不清楚,但我们可以从几个角度考虑其可能的原因。如果采用人工方法刺激含有单倍型胞核的整个卵子,使其单性发育,再假设在分化开始(器官形成)之前,该卵子具有与正常卵子相同的分裂次数,那么就细胞体积与其所含有的染色体数目之间的比例而言,它的每个细胞的体积势必会二倍于正常细胞。由于细胞是按照其基因来发育的,就有可能因为缺乏基因物质,不能对两倍大的胞质产生正常的影响。
另一方面,如果这类卵子在开始分化之前,比正常卵子多分裂一次,那么其染色体数目(即胞核体积)及其细胞体积就会维持正常的比例——整个胚胎也会含有二倍于正常胚胎的细胞,也会含有二倍的胞核。胚胎作为一个整体,将含有与正常胚胎相同的染色体数目。至于此例中较小的细胞体积,究竟会对发育过程产生何种影响,目前尚不明确。对单倍体细胞体积的观察,似乎表明虽然胞核是正常型的一半,但细胞体积却是正常的。看来,胚胎没有如前所述那样修正其胞核—胞质关系。
或许可以从另一方面考虑,是不是细胞体积正常而基因欠缺导致了人工培育单倍体的衰弱。含有一个精核的半个卵子,如果经历了与正常卵子相同的分裂次数,则胚胎细胞及其胞核之间将维持正常的体积比例。实际上,根据海胆胚胎的例子已得知了此类情况。它们发育成看起来正常的长腕幼虫,但没有任何一个度过了这一阶段,由于某种原因,甚至正常胚胎也很难在人工条件下培育到这一阶段之后。因此,目前尚不明确这些单倍体是否具备与正常胚胎相同的生活能力。Boveri和其他学者对海胆卵子切片进行了广泛的研究,多数切片可能小于半个卵子。Boveri认为,大多数单倍体在原肠形成时期之前或紧随原肠形成时期之后就已经夭折了。这些切片可能一直没能完全从手术中恢复,或者它们不具备胞质的全部重要要素。
如果将这些胚胎与正常二倍型卵子的裂球分离后所形成的胚胎进行比较,会发现一些引起人们兴趣的地方。当海胆卵子分裂为二个细胞、四个细胞或八个细胞时,用无钙海水对其进行处理,能够将裂球逐个分离。这里没有手术上的损伤,每个细胞都有双组的染色体。不过会有很多1/2裂球发育不正常,1/4裂球中能发育为长腕幼虫的更少,能度过原肠形成时期的1/8裂球可能一个也没有。这一证据表明,除染色体数目及胞核—胞质比例外,小体积本身也会产生不利影响。我们尚不清楚这意味着什么,但表面积和体积之间的关系随着细胞大小的变化而出现差异,这也许是产生上述结果的因素之一。
根据这些实验可知,没有多大把握能通过人工方法,减少已经适应了二倍型状态的物种中的卵子胞质,以获得正常的健壮的单倍体。不过,在自然条件下,存在若干单倍体的例子,其中一个例子中的二倍型物种的单倍体已经发育到了成年阶段。
Blakeslee在培育的曼陀罗中发现了一株单倍体(图10-2)。他细心地培养这株植物,把它嫁接到二倍体植物上,并保持了几年时间。这一植株除产生极少数单倍型花粉外,在全部主要性状上都与正常植株相近。其产生的花粉粒在极力度过成熟时期后,Blakeslee对其进行研究获得了一组染色体。

图10-2 曼陀罗的单倍体植株
一株曼陀罗单倍体植株(根据Blakeslee)
据Clausen和Mann(1924)的报告,烟草Nicotiana Tabacum与Nicotiana sylvestris杂交,产生两株各含有24条染色体的单倍型烟草,是Tabacum种的单倍数目。有一株单倍体相当于亲型Tabacum“变种”的“缩小版”,只是其性状表现得更为夸张,株高约为亲型的3/4、叶小、枝细,花也明显缩小一些。它不如亲型健壮、花朵繁盛,没有种子,花粉完全残缺。另一株单倍体相对于其Tabacum亲型变种,也呈现出同样的关系。以上两株单倍体的花粉母细胞经过不规则的第一次成熟分裂,少数或多数染色体进入两极,余下的留在纺锤体的赤道板上。第二次成熟分裂相对规则一些,但落后的染色体依然没有进入任何一极。
如果父母中的任一方是二倍体,那么该物种似乎自然能成功地得到少数单倍体。蜜蜂、黄蜂和蚁类的雄虫都是单倍体。蜂王的卵子含有16条染色体,接合之后得到8条二价染色体(图10-3)。在经历两次成熟分裂之后,又减为8条。卵子如果受精,就将发育成含有二倍数目染色体的雌性(蜂王或工蜂)。如果卵子不受精,就会按照一半的染色体数目进行单性发育。
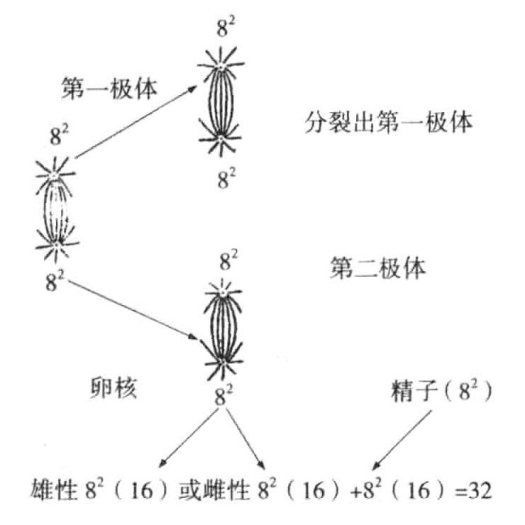
图10-3 蜜蜂卵子二次成熟分裂,卵子与精子受精
图示为蜜蜂卵子的二次成熟分裂,图下方表示卵子与精子受精后,染色体分裂为二,数目增加一倍
分别取雌雄两种蜜蜂的各种组织,对其胞核体积和细胞体积进行研究(Boveri、Mehling、Nachtsheim),发现二倍体与单倍体之间的差异通常不是一成不变的。不过,雌蜂和雄蜂在早期胚胎时期存在一种特殊现象,这一现象使问题更复杂一些。在雌雄蜜蜂的胚胎细胞中,似乎每一条染色体都分离为两部分,染色体的数目变成开始时的两倍。雌性胚胎细胞中也有同样的过程,甚至染色体还会重复这一过程,从而看起来具有32条染色体。这一证据似乎表明,染色体数目实际上并没有增加,只不过是各自“断裂”而已。如果这一解释成立的话,那么其体内基因数目也没有任何增加。雌蜂的染色体还是雄蜂的二倍。这一断裂行为与胞核大小之间究竟有何种关系(如果该关系真实存在的话),尚不明确。(https://www.daowen.com)
在雌雄蜜蜂的胚迹中,似乎没有发生断裂。如果该断裂已经发生,那么这些断片应该在成熟时期之前就已经重新结合了。
证明雄蜜蜂是单倍体,或至少其生殖细胞是单倍型的最佳证据是细胞在成熟时期的行为。第一次成熟分裂失败(图10-4a、图10-4b),一个不完全的纺锤体出现了,含有8条染色体。部分胞质分离出来,不含有染色体。第二纺锤体产生了,染色体各自分裂(图10-4d~g),这应该是纵裂方式的分裂,子染色体分别进入两极,各有一半。较大的细胞中分出一个小细胞,大细胞转变为含有单倍数目染色体的具备功能的精子。
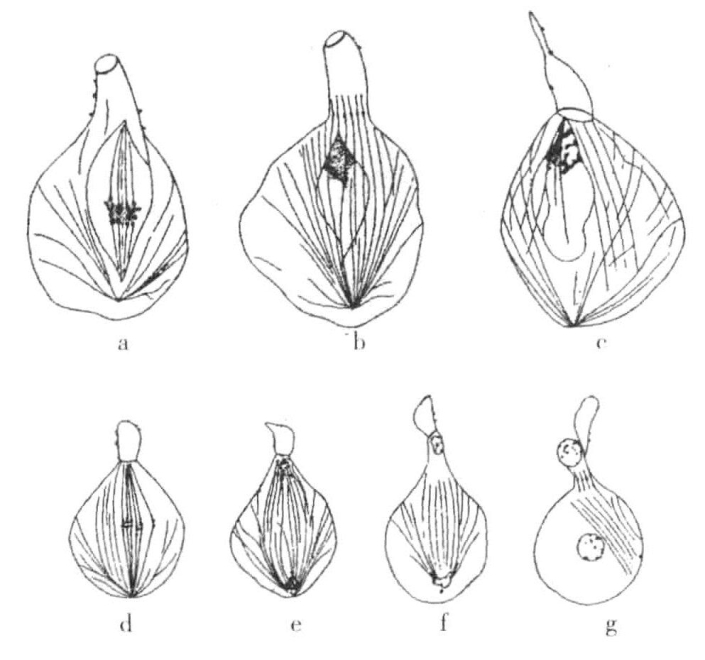
图10-4 蜜蜂生殖细胞的二次成熟分裂
雄性蜜蜂生殖细胞的二次成熟分裂(根据Meves)
锥轮虫Hydatina senta的雄虫是单倍体(图10-5c),雌虫是二倍体。在缺乏营养的情况下,或者以原生动物Polytoma为饲料时,只能产生雌性锥轮虫。雌虫是二倍体,其卵子在一开始也是二倍型。每个卵子只能分出一个极体——每条染色体分裂成相等的两半。从单性发育成雌虫的卵子,依然含有所有染色体。如果以其他食物(如眼虫)为饲料,就会产生一种新型雌虫。如果该虫刚从壳内孵出就与雄虫受精,则产生的卵子都是有性的,每个卵子放出两个极体,并维持单倍数目的染色体。已经进入卵子的精核与卵核结合,产生二倍体雌虫,重新建立一个单性繁殖谱系。但是,如果此类特殊雌虫不受精,就会产生比较小的卵子,该卵子将放出两个极体,大多保留了一半的染色体,在之后的单性发育过程中,产生雄性单倍体。雄虫则会在孵出几小时之后成熟,不再继续生长,并且在几日内死亡。
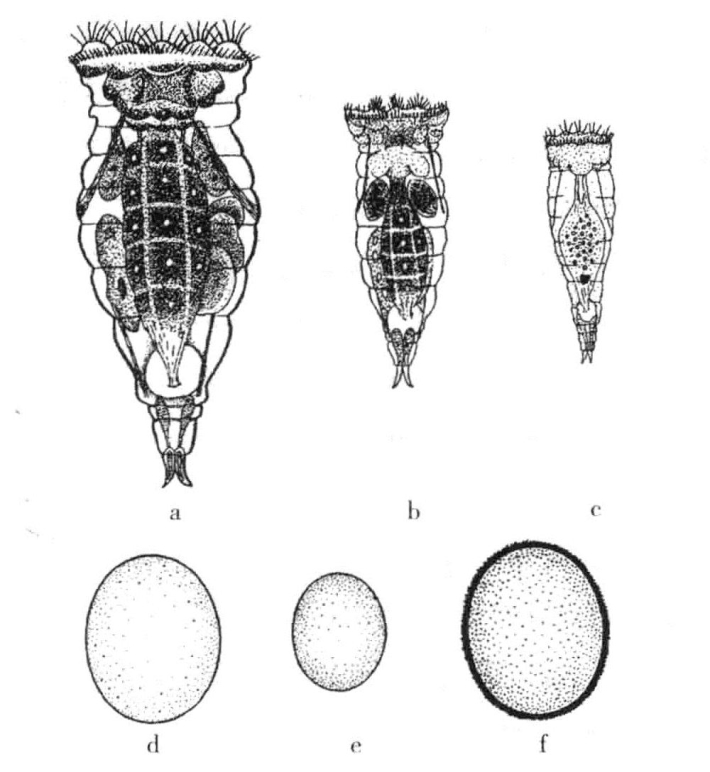
图10-5 锥轮虫Hydatina senta单性繁殖雌虫,有性繁殖雌虫与雄虫
a为锥轮虫Hydatina senta单性繁殖雌虫;b为同类幼年雌虫;c为同类雄虫;d为单性繁殖的卵子;e为发育成雄虫的卵子;f为冬季卵(根据Whitney)
Schrader证实雄性白蝇Trialeurodes vaporariorum是单倍体。A.W.Morrill发现,在美国没有交配的这类雌蝇只能产生雄性后代,在另一种同科生物中,Morrill和Back也发现了同样的现象。另外,Hargreaves和Williams相继提出报告,指出在英国没有交配的同种雌性白蝇只能产生雌虫。1920年,Schrader对美国种白蝇进行了研究,发现雌虫含有22条染色体,雄虫含有11条染色体。成熟卵子原本含有11条二价染色体,放出两个极体之后,卵子保留了11条单价染色体。卵子受精之后,增加了精核的11条染色体。没有受精的卵子进行单性发育,其胚胎细胞各含有11条染色体。雄虫生殖细胞成熟时,没有表现出任何减数分裂的迹象(甚至像蜜蜂那样的细微过程也没有),其均等分裂与精原细胞的分裂不存在差异。
正如Hindle的繁育实验所体现的,一些证据显示没有受精的虱卵可以发育成雄虱。一种恙虫Tetranychus bimaculatus的未受精卵子可以发育成雄虫,受精卵子可以发育成雌虫(根据一些观察报告得出)。Schrader(1923)证明单倍体雄虫只含有3条染色体,雌虫是含有6条染色体的二倍体。卵巢内的卵子最初含有6条染色体,然后,染色体两两接合,出现3条二价染色体。卵子受精时,增加了3条染色体,形成了雌虫的6条染色体。未受精的卵子直接发育成雄虫,其细胞内各含有3条染色体。
A.F.Shull对一种蓟马Anthothrips verbasci的雌虫进行了研究,发现没有交配时,未受精卵子只能发育成雄虫,这些雄虫大多是单倍体。
藓类和苔类的原丝体以及藓类的植株世代(配子体)都为单倍体。Wettstein用人工方法从原丝体细胞中得到二倍型原丝体和二倍型藓类植株。这一结果证明,这一世代与孢子体世代的区别不是由各世代的染色体的数目决定的,从某种意义上说,其只是一种发育现象,也就是说,孢子必定要经历配子体状态才能进入孢子体世代。