孟德尔两条定律的机制
在生殖细胞成熟末期,同样大小的染色体会接合在一起成对出现。这种情况会一直持续到生殖细胞的分裂,每组成对出现的两条染色体会分离开来,分别进入一个细胞。这样一来,每个成熟的生殖细胞就只会含有一组染色体(如图21和图22)。
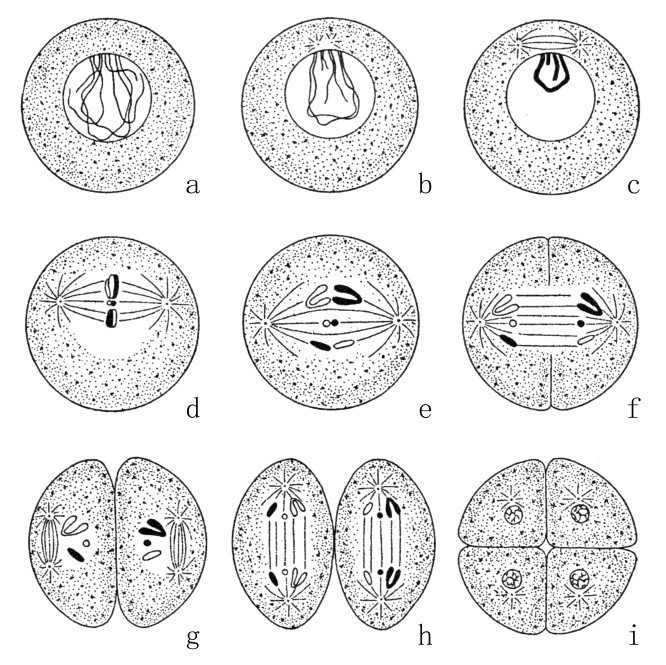
图21
精细胞的两次成熟分裂[4]。假定每个细胞含三组染色体,黑色代表来自父方,白色代表来自母方(a、b、c除外)。第一次成熟分裂是一次减数分裂,如图d、e、f。在第二次分裂或平均分裂中,每组染色体纵向地分裂成两条新的子染色体,如图g、h、i。
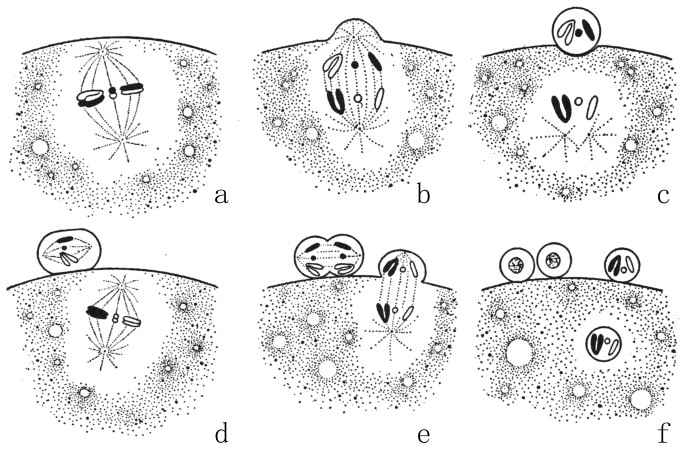
图22
卵细胞的两次成熟分裂。第一次形成纺锤体,如图a。来自父方和母方的染色体互相分离,如图b。图c已分裂出第一极体[6]。图d已形成第二纺锤体,且每条染色体纵向分裂成两半(均分)。图e中,已分裂出第二极体。图f所示为在卵细胞核中只留下一半的染色体(单倍)。
染色体在成熟时期的表现,与孟德尔第一定律相符。每对染色体中,来自父方的染色体与来自母方的染色体彼此分离,致使生殖细胞只含有每对染色体中的一条。当考虑到染色体都是成对存在时,我们或许会这样说,当生殖细胞成熟时,有一半的生殖细胞只含有每一对染色体中的某一条,另外一半的生殖细胞含有每一对染色体中的另一条。如果用孟德尔的单元概念去替换这里所说的染色体,其表述形式仍然是一样的。
一对染色体中的某一条是源于父方的,另外一条是源于母方的。当这种成对接合的染色体排列在纺锤体[5]上面时,如果来自父方的染色体去向一极,而来自母方的染色体去向另一极,那么所得的两个生殖细胞,将会分别与父母两方的生殖细胞相同。我们还没有先验的理由去假定,接合染色体会按这一路径行动,而且要证明它们不会按这一路径行动,也是极其困难的。因为从其本质来看,接合染色体在形状和大小上都是极其相似的,要将来自父方的染色体和来自母方的染色体分辨开来,几乎是做不到的。
不过,近几年我们发现,少数蚱蜢的某几对染色体之间,在形状上或依附于纺锤丝的方式上,有细微差异(如图23)。当生殖细胞成熟后,这些染色体先是两两成对接合,然后分离。由于它们保持着各自的差异,所以便于我们追踪它们前往两极的路径。(https://www.daowen.com)
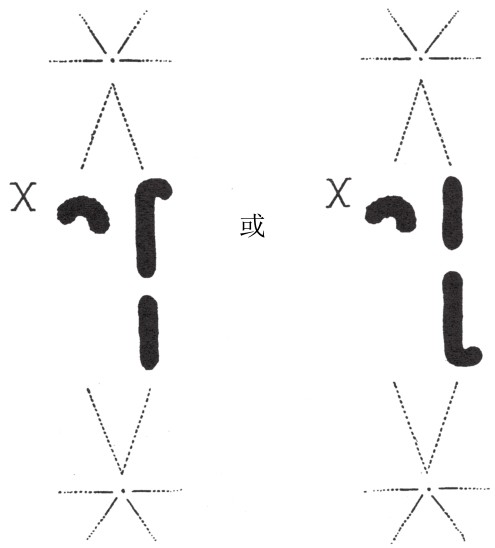
图23
X染色体与一组常染色体中的任意一条是随机自由组合的。
在这些蚱蜢中,雄蚱蜢存在某条无法组对的染色体,这条染色体与蚱蜢性别的决定有关(如图23)。当细胞成熟分裂时,这条无法组对的染色体会去向纺锤体的任意一极,且会成为其他成对染色体行动方向的“地标”。卡罗瑟斯(Carothers)女士是第一个观察到这一现象的人。她发现一曲一直的一对染色体,根据每一条染色体和性染色体的关系来看,这两条染色体分别去往哪一极是随机的。
继续深究,我们发现其他几对染色体中也出现了一些差异。研究这些成对染色体在成熟时的行动,我们再次发现,一对染色体分离后去往两极中的哪一极,同其他成对染色体的分布没有关系。从这里,我们得到了成对染色体分开后会与其他对染色体自由组合的客观证据。这一证据,也与孟德尔第二定律相符,该定律认为不同连锁群的基因是自由分配的。