多倍体小麦
在小型谷物的体细胞中存在多倍染色体,例如小麦、燕麦、黑麦和大麦。其中,对小麦系列谷物的研究最广泛,其杂交所得杂合子也都做过检查。单粒小麦[2]Triticum monococcum的染色体最少,只有14条(n=7)。单粒小麦属于Einkorn单粒群,据珀希瓦尔(Percival)(1921)所说,这种小麦能被追溯到新石器时代的欧洲。另外,二粒小麦[3]Emmer群有28条染色体,生长于史前时代的欧洲和公元前5400年的古埃及。直到希腊-罗马时代,该小麦才被含28条染色体的二粒小麦和含42条染色体的软粒小麦[4]取代(如图88)。在二粒小麦群组中,小麦的变种是最多的;在软粒小麦群组中,物种间的外形差异较大。
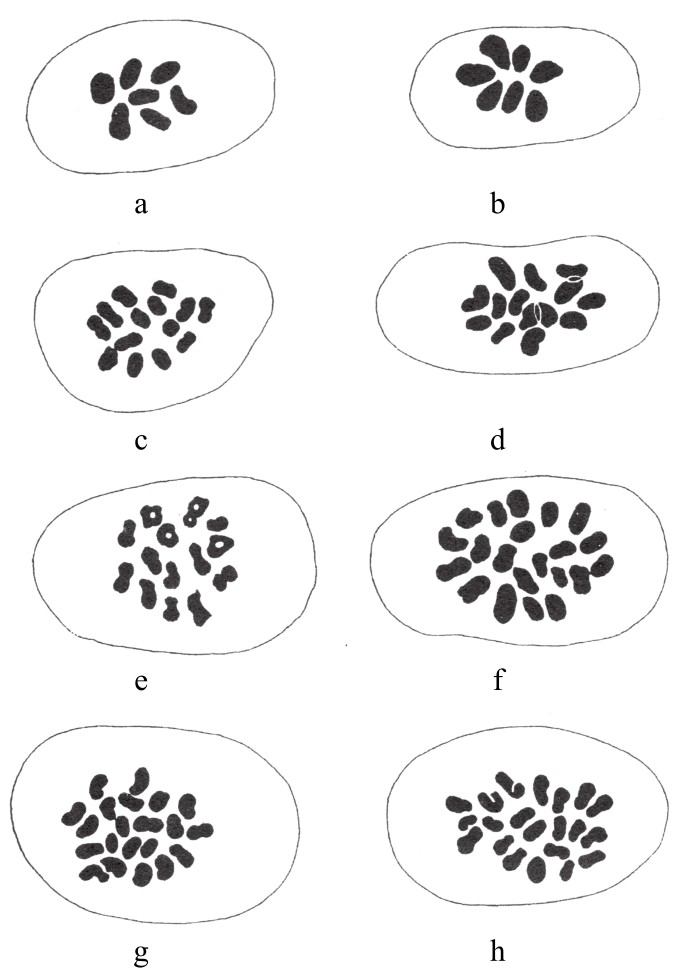
图88
二倍体、四倍体和六倍体小麦减数分裂后的染色体数目。
有几个学者研究过各类小麦的染色体。最近的研究有板村彻(Sakamura)(1920)、木原均(Hitoshi)(1919,1924)和萨克斯(Sax)(1922)的研究。下列数据大多数源于木原均的著作,一部分引用了萨克斯的论文。表4给出了观察所得的二倍体的染色体数目,以及观察所得或推测所得的单倍体的染色体数目。
表4 二倍体和单倍体小麦的染色体数目
单倍体群组由图88a(单粒小麦),图88c(坚粒小麦)和图88h(软粒小麦)表示。
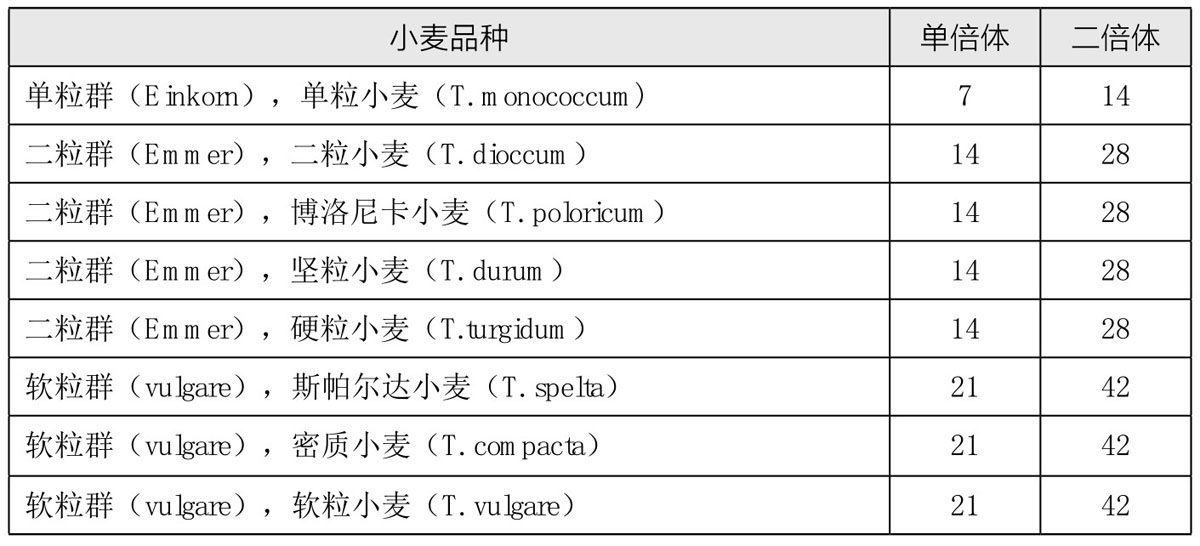
萨克斯用图89表示了这些群组各自的正常成熟分裂。单粒小麦的7条二价染色体(接合染色体)在第一次分裂中分离开来,每7条进入一极,且没有停滞的染色体。在第二次分裂时,第一次分裂而来的子细胞中的7条染色体中缢成两半,形成子染色体,每7条子染色体去向一极。二粒小麦在第一次成熟分裂时有14条二价染色体,每14条染色体去向一极。在第二次成熟分裂期,每条染色体中缢,在所得的子染色体中,每14条各自去向一极。软粒小麦在第一次成熟分裂期有21条二价染色体,每21条去向一极。在第二次成熟分裂期,每条染色体中缢,得到的子染色体每21条去向一极。
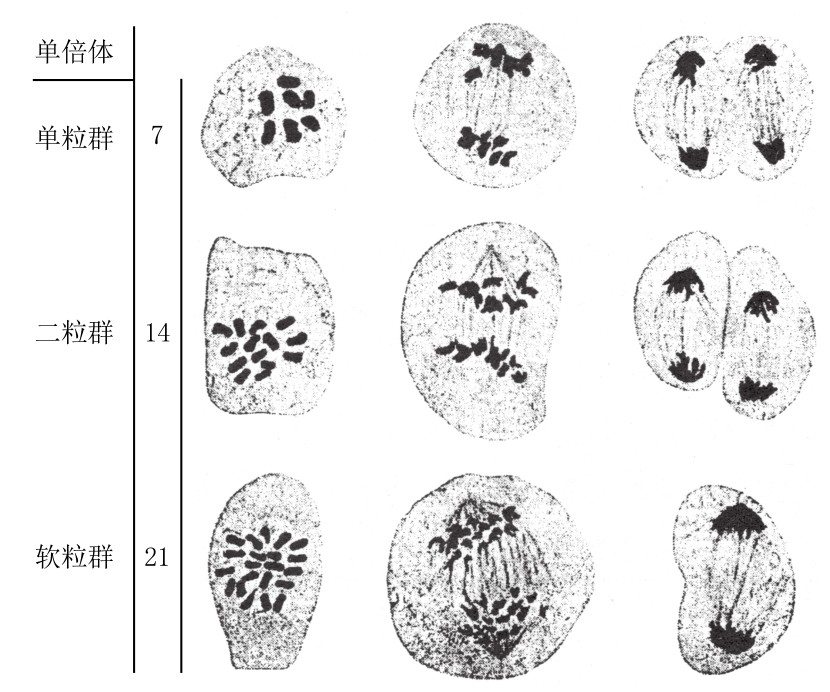
图89
二倍体、四倍体和六倍体小麦的第一次成熟分裂(减数分裂)。
这一系列类型可以解释为二倍体、四倍体和六倍体。每一种都是平衡的,也是稳定的。
将以上这几种拥有不同染色体数的品种进行两两杂交。在所得的后代中,会有一些生育能力较弱的杂合子,也会有一些完全不可育的杂合子。用于杂交的父方母方,有着不同的染色体数目,而这些染色体的行为可以揭示出一些重要的关系。下面几个例子可说明这一点。
木原均用有着28条染色体(n=14)的二粒小麦和有着42条染色体(n=21)的软粒小麦杂交,且对所得杂合子加以检测,他发现这类杂合子有35条染色体,因此判断它是一种五倍体的杂种小麦。它在成熟期(如图90a,图90b,图90c,图90d),会有14条二价染色体和7条单染色体。在第一次分裂时,二价染色体各自分裂后,每一极得14条。单染色体不规则地散落于纺锤体上,直到“减数”染色体分别进入两极,它们仍滞留于原地(如图90d)。接着,这些单染色体会中缢,其子染色体会进入两极,但运动方向十分不规则。当分布达到均匀之后,每极会获得21条染色体。
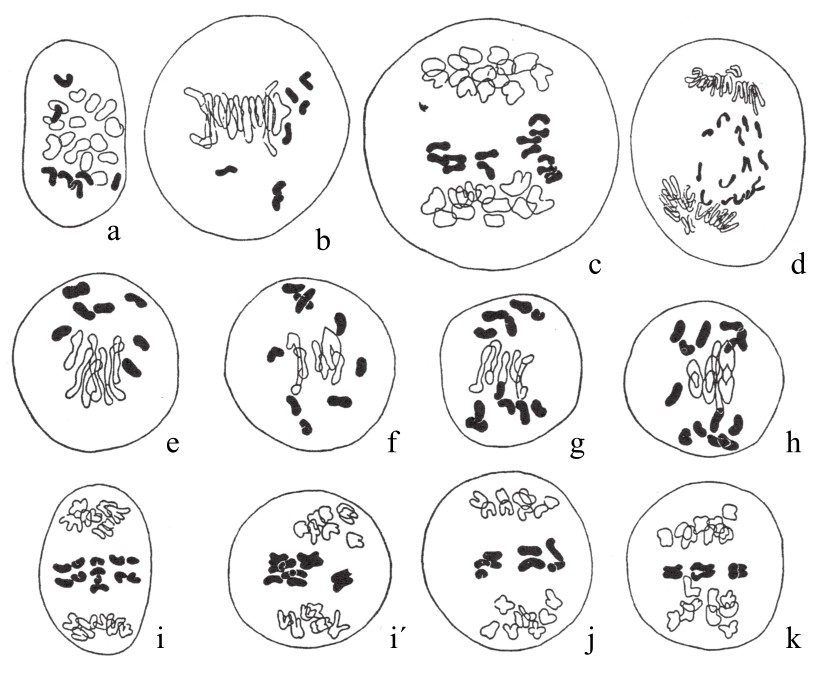
图90
杂合子小麦的减数分裂。
在此,我们应该顺便谈及萨克斯对三倍体小麦的研究结果:7条单染色体在此时并不分裂,而是不均匀地分布到两极,最常见的两极分配比例为3:4(如图91)。
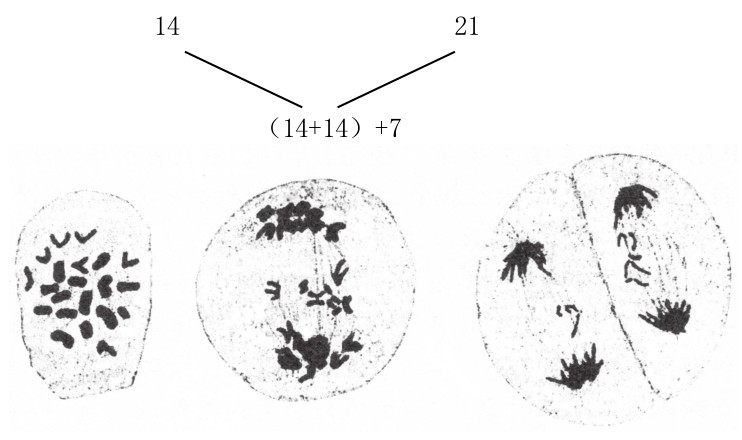
图91
二粒小麦和软粒小麦所得杂合子的减数分裂。
根据木原均的记载,在第二次减数分裂时,会有14条正在中缢的染色体和7条不会中缢的染色体。14条染色体分裂之后,两极分别有14条染色体,剩下7条单染色体随机分布——常见的是3条去向一极,4条去向另一极。但萨克斯认为,在第二次成熟分裂时,这7条染色体也会缢裂。
对于单染色体(两种解释都在其他生物中出现了先例),不管采用哪一种解释,有一个重要的事实都是很明显的,即仅在14条染色体间发生了接合现象。但发生接合的这14条染色体,究竟是由二粒小麦中的14条染色体和软粒小麦中的14条染色体接合而成,还是由二粒小麦中的14条接合成7条二价染色体,软粒小麦中的14条接合成7条二价染色体,共计达到14条二价染色体,然后在软粒小麦中还会余下7条单染色体,从细胞学的证据来看,现在还不明了。对这些物种进行遗传学方面的研究,抑或是对相似组合进行遗传学方面的研究,或许都能提供更有决定性的证据,但目前这样的研究还是很欠缺。
木原均又用有14条染色体(n=7)的单粒小麦Einkorn和有28条染色体(n=14)的二粒小麦Emmer杂交,所得杂合子是三倍体,有21条染色体。在此杂合子生殖细胞(花粉母细胞)成熟期间,其染色体分布的不规律性甚至超过了上一案例(如图90e,图90f,图90g,图90h,图90i,图90i´,图90j,图90k),接合在一起的染色体数目不恒定,且就算染色体间发生接合,也是不完全的。其二价染色体的数量多变,具体数目如表5所示:(https://www.daowen.com)
表5 二价染色体数目的变化

当第一次减数分裂发生时,二价染色体一分为二,所得两半各自进入一极。单染色体在进入两极中任意一极之前,不是每次都会发生分裂:有一些单染色体在未分裂的状态下就去向一极,而有一些单染色体在分裂后,其子染色体分为两半分别进入两极。7条单染色体停留在两极染色体群的中间平面上的情况,也并不少见(如图90i)。三次测定的数据如表6所示:
表6 染色体在两极的分布
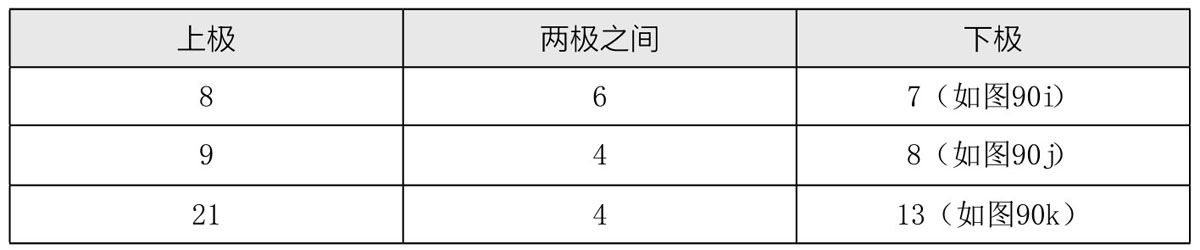
一般情况下,在第二次减数分裂期会有11条或12条染色体:一些是二价染色体(会发生中缢);另外一些仍旧是单染色体。前者(二价染色体)会正常分裂,其子染色体会分别去向两极;而后者(未发生缢裂的染色体)在没有缢裂的情况下分布于其中任意一极。
这一证据表明,要确定在杂合子中是哪些染色体接合到了一起是不大可能的(组成了二价染色体)。但因为二价染色体的数目未超过7,或许这可以解释为是亲代二粒小麦的14条染色体接合的结果。
在二粒小麦和软粒小麦杂交的少许案例中,我们得到了可育杂合子。木原均研究过子三代、子四代以及后面几代的杂合子,他观察了其染色体在成熟分裂期的行为。其后代各植株中染色体的数目是不一样的,而且一些染色体在成熟期的分布也是不规则的,这也导致了后期染色体的不规则分布,或者导致了子代重新形成如亲代那种稳定型,等等。这些结果,虽然对于杂合子的遗传研究是很重要的,但对于我们现在的研究目的而言,过于复杂了。
木原均还研究了软粒小麦和一种黑麦的杂合子,软粒小麦有42条染色体(n=21),黑麦有14条染色体(n=7),杂合子(有28条染色体)或许可称为四倍体。根据之前的观察,这两种亲代的差异较大,其杂合子是不育的,但其他观测者却认为是可育的。
从这个杂合子的生殖细胞形成到其成熟期,能观察到的接合染色体很少,甚至没有,如表7所示:
表7 生殖细胞成熟期的接合染色体数目
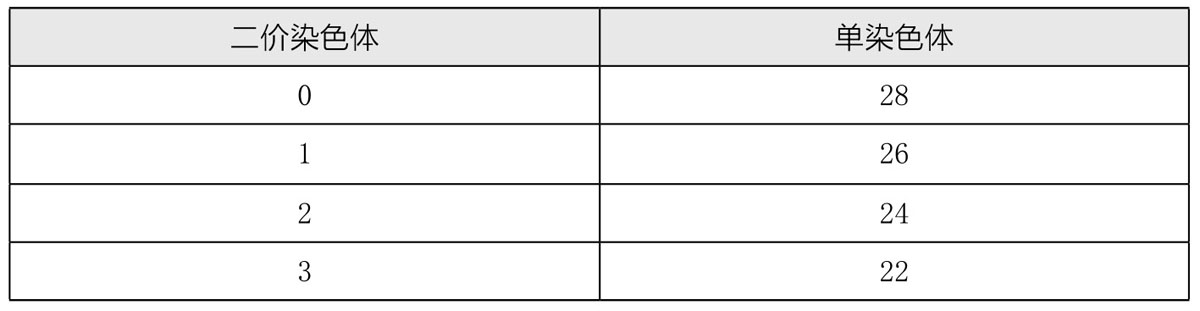
在第一次分裂中,染色体在两极的分布是极其不规则的,只有极个别染色体会在进入两极之前就分裂开来,部分单染色体会散落于细胞质之中。在第二次成熟分裂期,许多染色体会中缢开来,但在第一次成熟分裂时就已分裂的染色体,行动会比较迟缓,慢慢地去向一极;然而,落后的染色体数目远比第一次分裂时落后的染色体数目少得多。
在小麦和黑麦的杂交中,最有趣的一个杂交特征是杂合子几乎完全没有染色体的接合,这样就会导致染色体的不规则分布,而这种不规则分布,极有可能解释了杂合子不可育的原因。还有一种可能,即属于同一个物种的所有染色体(或大多数染色体)或许会去向另一极(发生的可能性极小),由此产生可孕性花粉粒。