复原突变(返祖性)对解释突变过程的重要性
一方面,如果隐性基因的产生是源于基因的缺失,那么,隐性纯种将不会再产生原有基因,要不然这就意味着,高度特化的某些物质竟能无中生有[9],这显然是说不通的。另一方面,如果突变是源于基因结构的改变,那么,不难想象有时突变基因可能会恢复原态。也许是因为对基因的了解甚少,我们对于这样一个论证,才不敢给出过高的评估。然而,用后一种观点来解释返祖突变体的出现,似乎较为合理。遗憾的是,关于这个问题的证据,尚不能完全令人满意。目前能确定的是,果蝇中存在的大量例子表明,在隐性突变的原种中,出现了与原种型[10]或野生型个体性状相同的个体。除非是在控制变量[11]的情况下,否则这类例子的出现还不能当成充足的证据,因为我们不能忽略野生型个体有可能会被隐性原种影响。然而,如果突变原种出现了多种突变性状,但有且只有一个性状发生复原,同时,还没有出现这些突变性状的其他组合形式,这种复原变化才是我们所需的证据。在我们培养的原种中,只出现了少数满足上述条件的案例,就其证据所涉及的范围来说,这也表明了复原突变这一现象是有可能发生的。同样,我们需要防止另外一种可能性的发生,那就是一段时间之后,有些突变原种,看起来像是或多或少地失去了原种的一些性状,然而,当将其杂交之后,这些失去的突变性状却又恢复了。例如,第四染色体上的弯翅性状,其本身就是不稳定的,极易受环境影响,如果不对培育的个体加以选择,其性状便会复原到原种的样子。如果将这种个体与野生原型杂交,再将所得子一代自交,那么所得的弯翅后代中,预计很多个体会重现弯翅性状。在另一个被称为盾片果蝇的突变原种中,也出现了类似结果。所谓盾片性状,即果蝇的胸膛缺少了某些刚毛。在某些纯种盾片原种个体中,出现了“失去”刚毛的现象。表面上,这些突变体中,已有复原回野生型的个体。但通过将这些个体与野生型原种交配,却并未出现复原的野生型个体。在子二代中,盾片果蝇再次出现。对这个例子的研究表明,果蝇恢复正常,是隐性基因发挥作用的结果,因为当其出现在盾片原种的纯合状态下时,“失去”的刚毛会再度长出。以上实验结果除了与正在讨论的问题有所关联之外,这个事实本身也是有趣而重要的,即一个隐性基因的出现会使得突变性状回到原种型性状。
最后,还存在一种独特的复原现象,即细眼这一显性或半显性性状可以复原到正常眼(如图49a、图49b)。根据梅(May)和泽莱尼(Zeleny)多年的观察得知,细眼会恢复到正常眼,这个证据可以用来充当复原突变发生的证据。复原突变发生的频率因原种的不同而不同,据估计,1 600次变异中大约会发生1次复原变异。之后,斯特蒂文特(A. H. Sturtevant)和摩尔根发现,当细眼基因复原发生之际,在细眼基因的附近会发生基因的交换。斯特蒂文特在确定所发生的变化的性质方面,找到了关键证据。
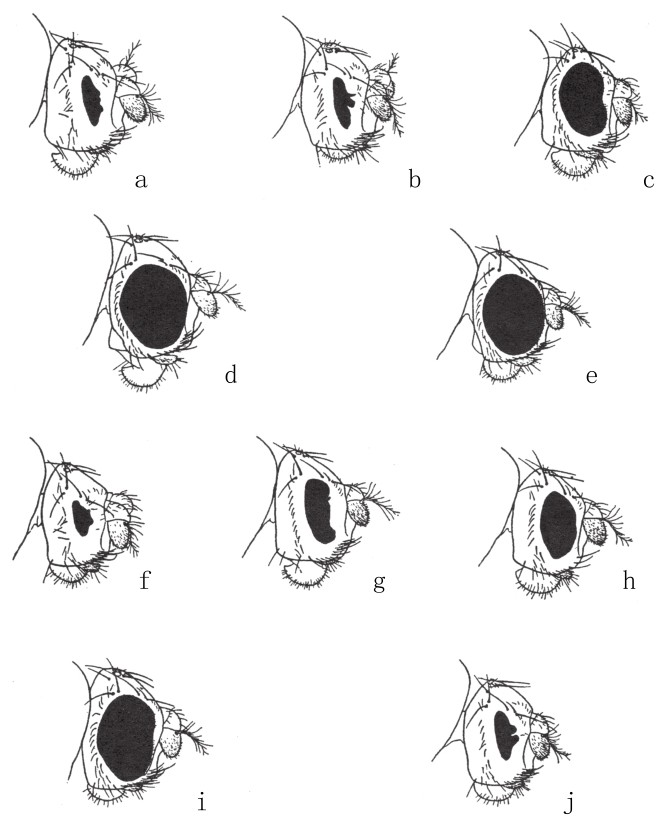
图49
各种细眼类型果蝇的眼睛。图a为纯合子细眼雌蝇;图b为细眼雄蝇;图c为细眼对圆眼雄蝇;图d为复原所得的纯合子圆眼雌蝇;图e为复原所得的携带圆眼基因的雄蝇;图f为双细眼雄蝇;图g为纯合子次细眼雌蝇;图h为次细眼雄蝇;图i为次细眼对圆眼雌蝇;图j为双次细眼雌蝇。
证明每次复原都会发生基因交换的方法如下:在细眼基因的左端(相距0.2个单位处)有一个叉毛基因,在细眼基因的右端(2.5个单位处)有一个合脉基因。一只雌蝇有如下组合:一条X染色体上含前述三个基因,细眼基因位于叉毛基因与合脉基因之间;另一条X染色体上不仅有细眼基因,还有叉毛基因与合脉基因的野生型等位基因(如图50)。将这样的一只雌蝇与叉毛、细眼、合脉雄蝇交配,所得子一代中的普通雄蝇,接受了母方的一条含有叉毛、细眼、合脉的X染色体,或是另一条含有非叉毛、细眼、非合脉的X染色体,其表现出的性状,要么是叉毛、细眼、合脉,要么是细眼。虽然复原突变很少发生,但当其发生时,即当一只圆眼雄蝇出现时,我们就会看到在叉毛基因和合脉基因之间发生了交换。例如,复原所得的雄蝇不是叉毛就是合脉,从不会出现兼具叉毛与合脉的性状,也不会出现兼具非叉毛和非合脉的性状。所以在母体染色体内,在叉毛基因和合脉基因之间一定发生了交换。叉毛基因和合脉基因之间的交换,总计不超过所有交换基因的3%,其中包含了所有的复原突变基因。
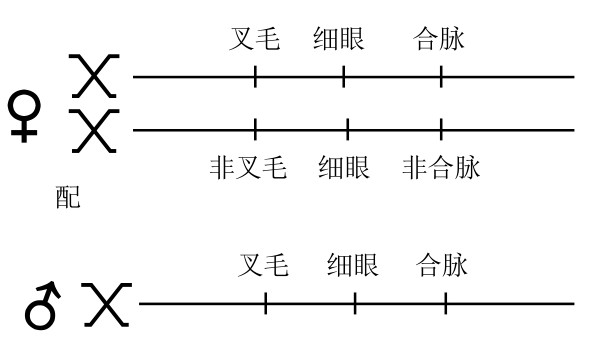
图50
一只细眼雌蝇在叉毛、合脉上为杂合子,该雌蝇与一只叉毛、细眼、合脉的雄蝇杂交。
为了简化表述,前面只谈及了子代中雄蝇的复原情况。当然,复原的染色体也可能传递给卵细胞,从而发育出雌蝇。我们可以设计这样一个实验,以便在子代雌蝇中检验交换的证据。子代的普通型雌蝇尽是纯合子的细眼基因(如图49a),而复原型雌蝇会有杂合子的细眼基因,再加上叉毛基因,或者加上合脉基因,没有一只是兼具叉毛基因和合脉基因的个体,也没有一只是兼具非叉毛基因和非合脉基因的个体。
引起复原圆眼的交换作用,不仅会使一条X染色体失去一个细眼基因,还会将这个细眼基因转移到另一条含有细眼基因的X染色体上[12](如图51a)。带两个细眼基因的雄蝇和带一个细眼基因的雄蝇,在外形上很相似,但前者眼睛更小,其小眼的数目也比较少。前者被称为双细眼(如图49f)。至今为止,还没有人观察到在任何其他突变中有两个等位基因出现在同一直线序列上的情况。作图时,我们只能假设在交换前,两个细眼基因是相对的,而在交换时,却稍微移动了一下位置。结果,双细眼雄蝇的染色体至少延长了一个细眼基因的长度,与之相反的是,另外一条染色体的长度则因为减少了一个细眼基因而缩短了。
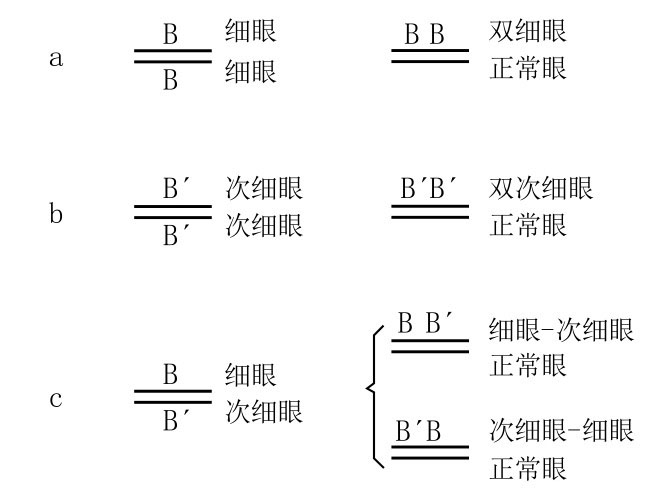
图51
细眼、次细眼和细眼-次细眼的突变。
斯特蒂文特对该复原理论做过一些关键性的测验。细眼基因的等位基因(由细眼基因突变而来)被称为次细眼基因(如图49g和图49h),次细眼型与细眼型在两眼的大小以及小眼的数量上有些许不同。次细眼原种会发生复原突变(如图51b),产生一个与野生型非常相似的完全圆眼型果蝇及一个被称为双次细眼[13]的新型果蝇(如图49j)。
当复原突变发生时,一条染色体上带细眼基因,另一条染色体上带次细眼基因(如图51c)的雌蝇会产生完全圆眼野生型果蝇和细眼-次细眼型果蝇,或者是完全圆眼野生型果蝇和次细眼-细眼型果蝇(如图51c)。(https://www.daowen.com)
斯特蒂文特还利用了细眼-次细眼型和次细眼-细眼型来证明如下事实:假设突变基因都位于同一条染色体,那么当细眼-次细眼型果蝇与正常型果蝇发生交换时(如图52a),结果所得要么是细眼叉毛型果蝇,要么是次细眼合脉型果蝇;当次细眼-细眼型果蝇和正常型果蝇发生交换时(如图52b),结果要么是次细眼叉毛型果蝇,要么是细眼合脉型果蝇。
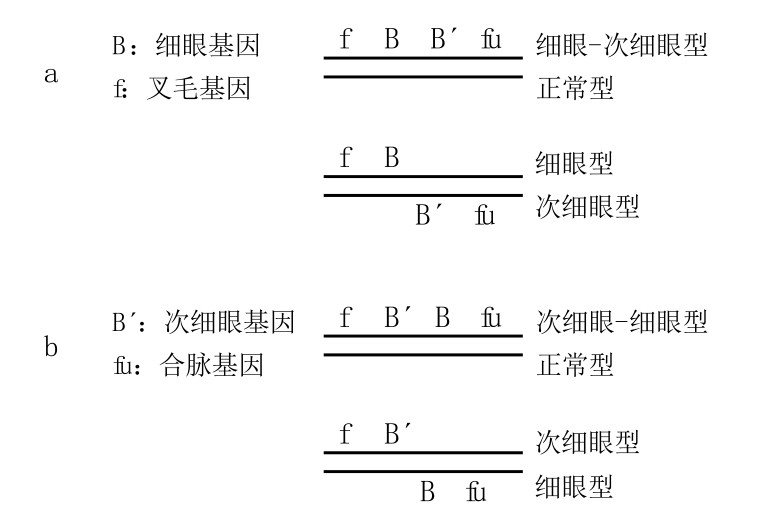
图52
图a是细眼叉毛果蝇到非细眼合脉果蝇的突变;图b是非细眼叉毛果蝇到细眼合脉果蝇的突变。
因此,这两型中的各个基因不仅保留了各自的特性,而且也维持着基因之间的顺序。从fBB´fu和fB´Bfu的组合方式来看,基因的排列顺序是一目了然的。而且,在所有例子中,B和B´之间的断裂,都与原来的基因顺序相吻合。
这些结果,为细眼型果蝇是因为交换而复原的说法提供了重要的证据。目前,这是唯一的案例。在X染色体上的细眼基因位上,似乎有一种特性,这种特性促使等位基因之间发生交换。斯特蒂文特将其称为“不等交换”[14]。
这一结果也引出了这样一个问题:是否所有的突变都是源于交换?在果蝇实验中,我们所得到的确切证据表明,基因交换不能普遍解释为突变。其中的一个原因是我们都知道的:突变既可以发生于雌蝇身上,也可以发生于雄蝇身上,而雄蝇不存在X染色体的交换。