鸟型(WZ-ZZ)
性别决定机制的另外一种类型是鸟型,如图118所示。雄鸟有两条相同的性染色体ZZ。它们会在一次成熟分裂期分开,致使每一个成熟的精细胞都含有一条Z染色体。雌鸟有一条Z染色体和一条W染色体。当卵细胞成熟(分裂)之后,每个分裂所得的卵细胞会得到一条染色体。这样一来,一半的子卵细胞含有Z染色体,另一半的子卵细胞含有W染色体。任何含有一条W染色体的卵细胞和含有Z染色体的精子结合会得到雌鸟(WZ),任何含有一条Z染色体的卵细胞和含有Z染色体的精子结合会得到雄鸟(ZZ)。
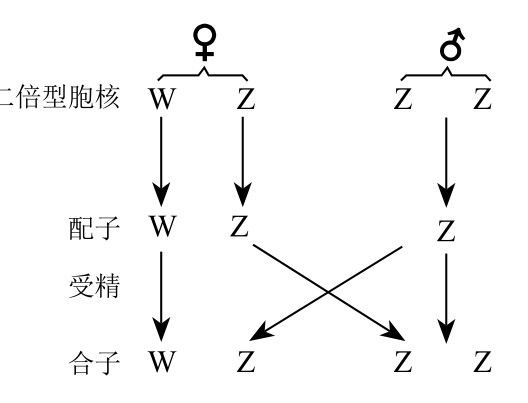
图118
WZ-ZZ型性别决定机制。
在此,我们再次发现了一种可以自动得出相同数目的雌雄两种个体的机制。同之前的例子相同,受精时期发生染色体接合会得到1:1的雌雄比例。鸟类的这种遗传机制的证据来源于细胞学和遗传学两个方面,但是细胞学证据目前还不能完全令人信服。
斯蒂文斯(Stevens)表示,公鸡有两条同样大的长染色体,假定为ZZ[4](如图119),而母鸡只有一条这样的长染色体。石和后(Shiwago)和汉斯证实过这样的关系。
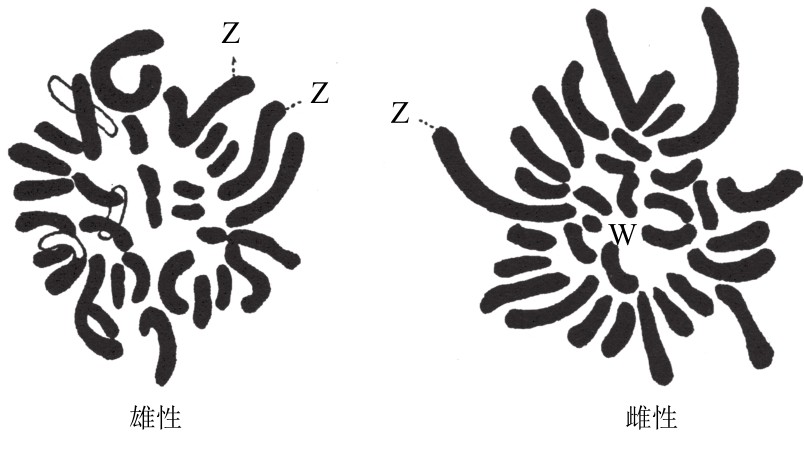
图119
公鸡和母鸡的染色体群。
鸟类的遗传学证据是毋庸置疑的,这一证据源于性连锁遗传。如果让黑色狼山型公鸡和花纹Plymouth rock型母鸡交配,所得雄性个体都有花纹,而雌性个体都是黑色的(如图120)。如果Z染色体携带分化基因,则预期所得结果应和上述相同,因为子一代的雌性会从其父代得到Z染色体。如果让子代的后代杂交,会得到花纹公鸡、花纹母鸡、黑色公鸡、黑色母鸡共四种类型,比例为1:1:1:1。
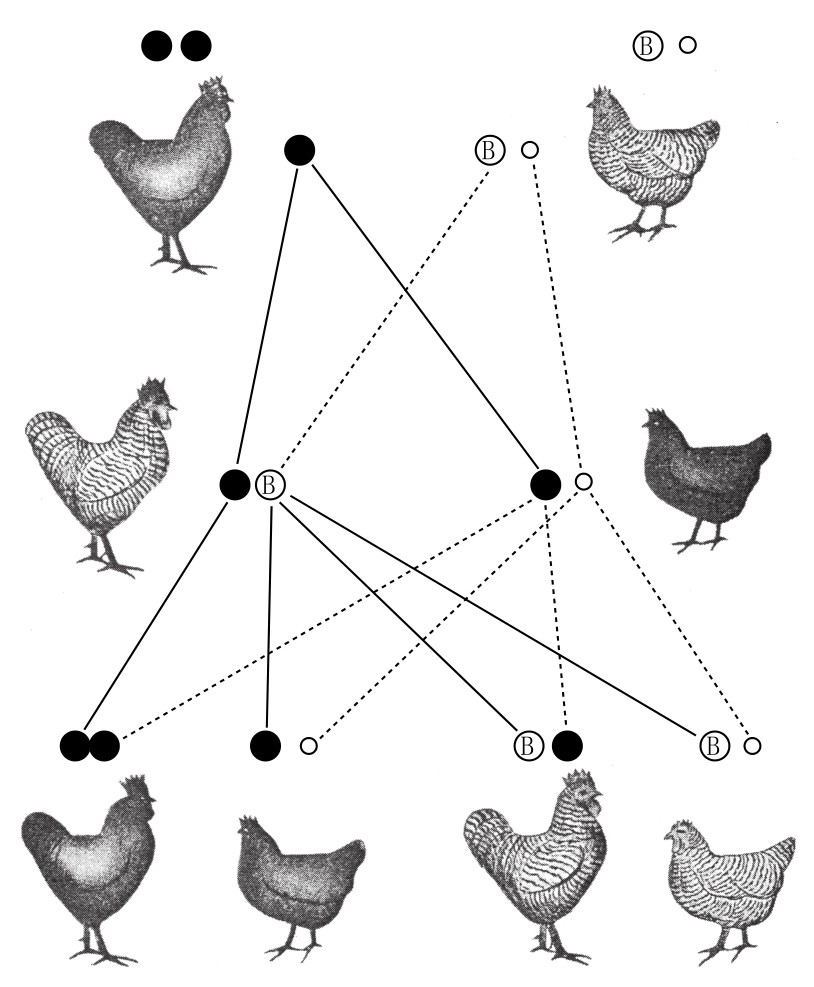
图120
带花纹的家禽与黑色家禽的杂交,体现了性连锁型遗传规律。
在蛾类中也发现了同样的遗传机制,其细胞学证据更为明确。当黑色醋栗蛾的雌性野生变体型与浅色的雄性突变型交配时,其雌性子一代和父方比较相似,拥有浅色性状;而其雄性子一代则和母方比较相似,拥有黑色性状(如图121)。子一代雌性从父方那里得到一条Z染色体;子一代雄性从母方那里得到一条Z染色体,从父方那里得到另外一条Z染色体。母方的Z染色体携带显性的黑色基因,因此其雄性子一代都是黑色性状。
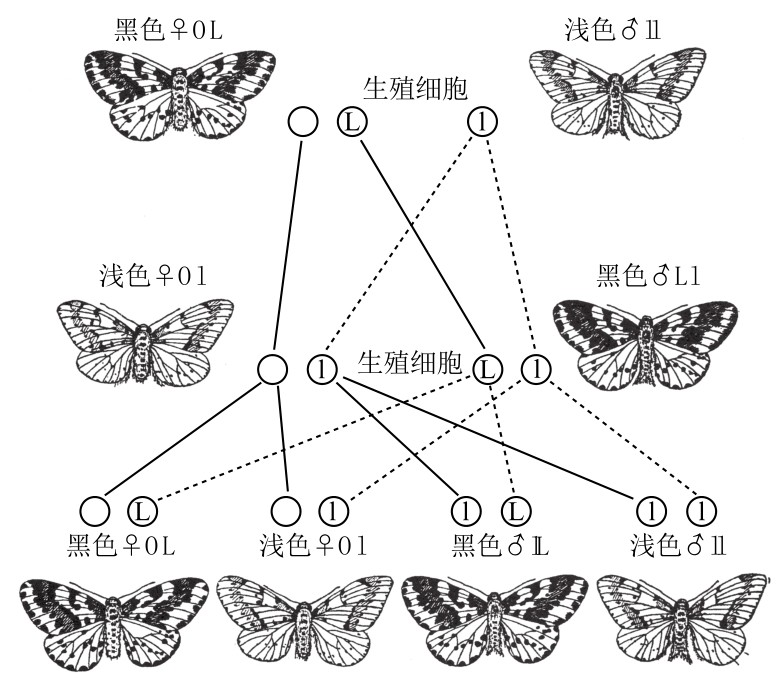
图121
醋栗蛾的性连锁遗传。
田中义麿(Tanaka Yoshimaro,1884—1972)发现,蚕蛾幼虫时期的透明质皮肤是一种性连锁性状,这种性状似乎也是通过Z染色体遗传给下一代的。
Fumea casta型雌蛾有61条染色体,雄蛾有62条染色体。卵细胞中的染色体接合之后,会有31条(图122a)。在第一次分裂出极体时,30条二价染色体分离后分别进入相反的两极;第31条单价染色体在未分裂的情况下随机进入某极(如图122b和b´)。结果,有半数卵细胞含有31条染色体,半数卵细胞含有30条染色体。在第二次分裂出极体时,所有染色体一分为二,所得子细胞中染色体的数目和分裂之前的数目相同(即30或31)。当精细胞成熟时,染色体会接合成31条二价染色体。在细胞的第一次成熟分裂期,每组染色体(即二价染色体)会分离开来;在细胞的第二次成熟分裂期,分离而来的染色体又会一分为二,每个精子都会含有31条染色体。卵细胞受精后可得到下列组合:
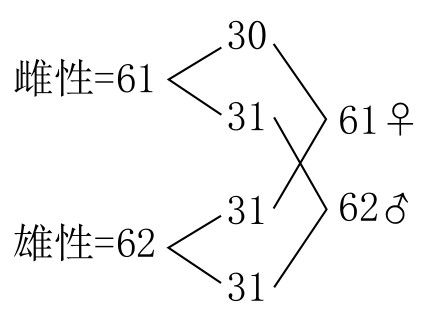
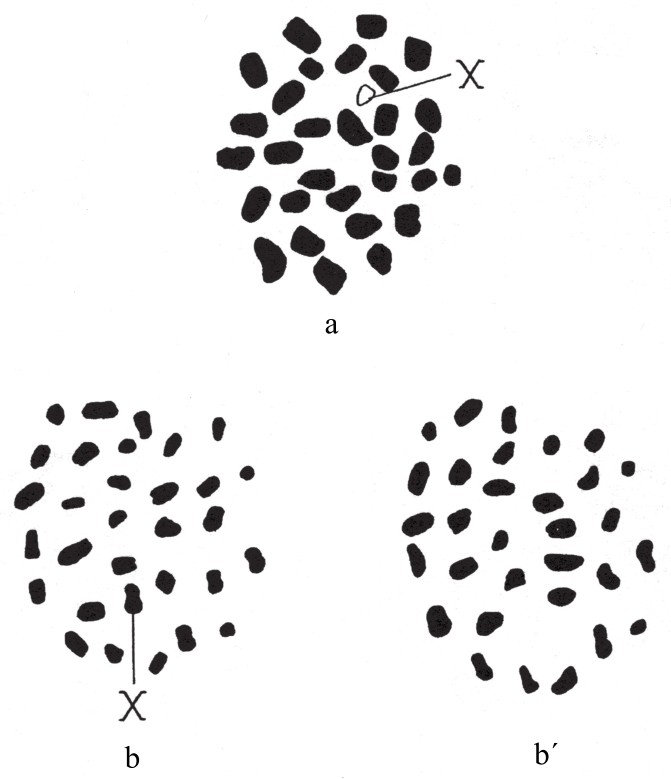
图122
图a为Fumea casta型雌蛾的卵细胞减数分裂后的染色体群;图b和图b´为卵细胞第一次成熟分裂时两个极体的染色体群。单条X染色体只存在于某一极。
赛雷尔发现,Talaeporia tubulosa型雌蛾有59条染色体,雄蛾有60条染色体。但在Solenobia pineti型雌蛾或雄蛾以及其他几种蛾类中,均看不到未成对的染色体。另一方面,Phragmatobia fuliginosa型蛾却有一条或两条包括性染色体在内的复染色体[5]:雄蛾含有两条这样的复染色体;雌蛾只含一条与雄性的复染色体相似的染色体。在含有W因子和Z因子不分开的染色体的其他蛾类中,这样的现象似乎也出现过。
费德莱让Pygaera anachoreta型和Pygaera curtula型这两种蛾杂交,证明了蛾类的性连锁遗传。这一案例很有趣,因为每一物种的雌雄幼虫都是相似的,但不同物种的幼虫会表现出一些物种间的差异。在同一物种内没有二形的种间差异[6],成了子代幼虫性二形的基础(当杂交依循“一个方向”进行时),其原因正如结果所示,两种幼虫间基因的主要不同在于Z染色体。当anachoreta蛾为母方,而curtula蛾为父方时,所得杂合子幼虫在第一次蜕变之后,就会变得极不相同:杂合子中的雄蛾幼虫会与母族(anachoreta)的性状十分相似,而杂合子中的雌蛾幼虫会与父族(curtula)的性状非常相近。
若反着来交配,即anachoreta蛾为父方,curtula蛾为母方,所得的子代杂交种则完全相似。这一结果,可以用下列的假设来解释:anachoreta蛾的Z染色体所携带的一个(或多个)基因,相对于curtula蛾的Z染色体上所携带的一个(或多个)基因,呈显性。这一案例的有趣之处在于,某物种的基因相对于另一物种的相同染色体上的等位基因来说,呈显性。这种解释,同样适用于由子代雄蛾与任一亲代回交所得的孙代,但需要将后代的三倍性考虑在内(见第九章)。
我们没有理由去假设XX-XY型的性染色体和WZ-ZZ型的性染色体是相同的。反之,我们也无法想象,某一物种直接变成另一物种。尽管两种类型中所包含的基因是相同的或几乎相同的,但决定两性平衡的某种变化,依旧可以在两种类型中独立发生——这一假设在理论上是不存在困难的。