第十章 单倍体
第十章 单倍体
遗传学证据表明,生物的正常发育至少需要一组完整的染色体。只有一组染色体的细胞被称为“单倍型”(haploid)细胞;由多个单倍型细胞组成的个体,有时被称为“单倍体”(haplont)。胚胎学方面的证据同样表明,一组染色体对于生物的发育是很有必要的。然而,并不能据此断言,就所涉及的发育条件来说,单倍体能直接取代二倍体,而不会有任何严重的后果。
遗传学证据表明,生物的正常发育至少需要一组完整的染色体。只有一组染色体的细胞被称为“单倍型”(haploid)细胞;由多个单倍型细胞组成的个体,有时被称为“单倍体”(haplont)[1]。胚胎学方面的证据同样表明,一组染色体对于生物的发育是很有必要的。然而,并不能据此断言,就所涉及的发育条件来说,单倍体能直接取代二倍体,而不会有任何严重的后果。
接受人工刺激之后,卵子会发育成只有一组染色体的胚胎。但卵细胞在发育之前,其细胞内的染色体加倍(抑制细胞质的分裂,使染色体加倍而细胞不分裂)的情况也不在少数,而且染色体加倍后比单倍体存活得更好。
切下海胆卵细胞上的一小片,使其与单倍型精子结合,我们能得到一个源自父代单组染色体的胚胎。在蝾螈受精之后,斯佩曼(Spemann)和巴尔策(Baltzer)马上缢裂蝾螈的卵细胞,分离出一片只含单精核的卵子细胞质(如图83)。而且,巴尔策随后将其中一个胚胎培育到了变态[2]时期。
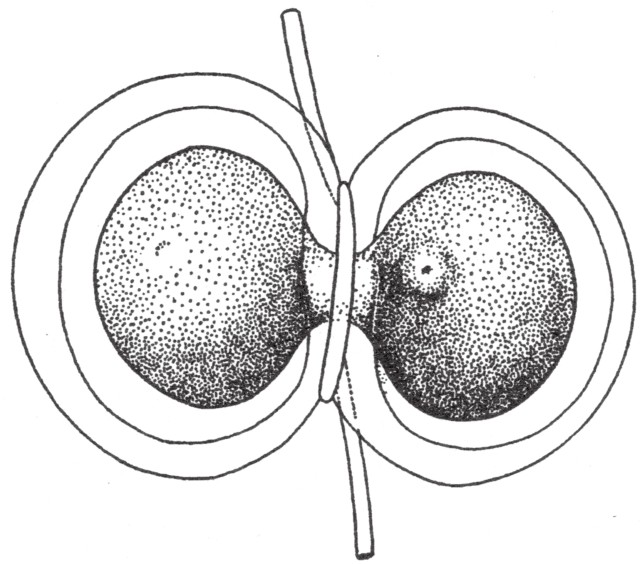
图83
蝾螈卵细胞在受精之后立即缢成两部分。右边所示为极体。
如果长时间用X射线或激光照射蛙的卵细胞,损伤或破坏其染色体,像奥斯卡(Oscar)和赫特维西(G. Hertwig)所展示的那样,使这些卵细胞受精,那么,在由这些受精卵所得到的胚胎中,只有其原型一半的染色体数。相反,如果长时间照射精细胞,它们或许能够进入卵细胞,但对发育不再起作用。在如此情境下,卵细胞只有卵核的单组染色体,可能继续发育一段时间。也就是说,卵核中的染色体能够分裂,但细胞质不能分裂,从而在开始发育之前,卵细胞又恢复了完整数目的染色体。
经由上述途径得到的人工单倍体,多数是很虚弱的。在大多数案例中,人工单倍体会在未成年前死亡。其死亡原因还不是很明确,但我们可以思考以下几种可能性。若用人工手段去刺激含单倍型胞核的卵细胞,使其进行单性发育,加之,如果在细胞分化形成器官之前,该卵细胞的分裂次数和正常卵细胞的分裂次数相等,那么,从细胞体积与染色体数目之间的比例维持正常这一点来看,每个细胞的体积会是正常细胞体积的两倍。从细胞发育需要依靠其基因这一点来看,其体积两倍于正常细胞的细胞质不足以产生正常效应,这是因为基因物质欠缺。
另一方面,如果在这种卵细胞开始分化[3]之前(器官形成之前),该卵细胞的分裂次数比正常卵细胞的分裂次数多一次,那么染色体数目(细胞核体积)和细胞体积的比例就会是正常的——在整个胚胎中,其细胞和细胞核是正常胚胎的两倍。胚胎作为一个整体来看,会拥有与正常胚胎相同的染色体数。在这样的案例中,较小体积的细胞,其体积究竟要比正常细胞体积小多少,才会对其发育造成影响,我们目前尚不清楚。对这些单倍体生物的细胞体积加以观察,我们发现:似乎单倍体细胞有着正常的体积,但其细胞核只有正常细胞核的一半大小。这样说来,胚胎并未像前面所说的那样校正细胞核与细胞质之间的关系。
或许我们还有另外一种方法去判断,人工单倍体的衰弱是不是由于人工单倍体细胞与正常细胞一样大,但其基因数量却没有正常细胞的多。如果含一个精核的半个卵细胞能和正常卵细胞一样经历两次分裂,且分裂特征相同,那么,胚胎细胞和细胞核之间将维持正常的体积比例。事实上,这种类型的海胆胚胎很早就存在。这些海胆,着力想要变成看起来正常的长腕幼虫,但没有一只能挺过幼虫时期。出于某种因素,即使是正常胚胎,想要在人工条件下活过这段时期也是很艰难的。因此,这些单倍体是否能和正常胚胎一样存活下来,不能确定。博维里(Boveri)和其他学者对海胆卵细胞的切片做了广泛研究。他的结论是,绝大多数单倍体海胆在原肠形成时期或是其后不久都会死亡。这有可能是因为这些片段并未从切片这一操作中恢复过来,又或者是因为这些片段不含细胞质的所有重要成分。
将这些胚胎与从正常型二倍体卵裂球[4]中分离出来形成的胚胎做比较,是很有趣的一点。用以下方法分离胚卵裂球是可行的,即当海胆卵细胞一分为二,二分为四,四分为八时,用无钙海水处理,可将这些卵裂球分离出来。这一操作并不会伤害卵裂球,每个分裂所得的细胞都含有双组染色体。然而,有许多1/2卵裂球发育不正常,1/4卵裂球中能发育到幼虫时期的更少,能挺过原肠形成时期的1/8卵裂球可能一个也没有。这一证据表明,除了染色体数目和细胞核——细胞质的比例,细胞自身体积较小对个体发育也会有不利影响。这意味着什么,我们暂时还不知晓,但细胞体积和表面积的关系,会随着细胞大小的变化而有所变化,这或许是上述结果的一个影响因素。
从这些实验来看,在已经适应二倍体状态的物种中,若要用人工方法减少卵细胞的细胞质来获得有活力的正常单倍体,希望不大。然而,在自然条件下,存在若干单倍体的案例,其中有一个二倍体物种的单倍体存活到了成年。
布莱克斯利在栽培曼陀罗时,发现了一株单倍体(如图84)。他将其嫁接于一株二倍体植株上,并悉心照料,使得这株单倍体存活了好多年。这株植物,除了会产生少量的单倍体花粉之外,其他主要性状都和正常植株的相似。这些花粉,都是在奋力拼搏度过成熟期之后,才得到了一组染色体的。

图84
曼陀罗的单倍体植株。
根据克拉苏森(Clasusen)和曼(Mann)在1924年的研究,在tabacum烟草和sylvestris烟草的杂交过程中,出现过两株单倍体烟草植物。每一株单倍体植株都含有24条染色体,这是tabacum烟草品种中的单倍数目。这两株中有一株,是亲代植株tabacum“缩小版”的“变体”,但其在性状的表达上,它略显夸张。其高度是亲代的3/4,叶子比亲代小,枝干也比较细,花朵明显比亲代小一些。该株烟草并没有亲代那么健壮、花朵茂密,且不结果,其花粉也是完全残缺的。前面提到的另外一株单倍体烟草,与它的tabacum亲型变种之间,也有相似的关系。这两株单倍体的花粉母细胞,经历了不规律的第一次成熟分裂时期,出现极多或极少的染色体去向两极,剩下的染色体则留在纺锤体的赤道面上。第二次成熟分裂会稍微规律一点,但停滞的染色体仍然不能进入任何一极。
在亲代中只有一方为二倍体的物种中,自然界似乎创造出了少数的单倍体。蜜蜂、黄蜂和蚁类的雄虫都是单倍体动物。蜂后的卵细胞含有“16条染色体”,在与精子中的染色体接合后会变成8条二价染色体(如图85)。两次成熟分裂之后,每个卵细胞含有“8条染色体”[5]。如果卵细胞受精,那么会得到含二倍染色体数目的雌蜂(蜂后或工蜂);如果卵细胞未受精,那么这个卵细胞会继续进行单性发育,最终发育成雄峰。
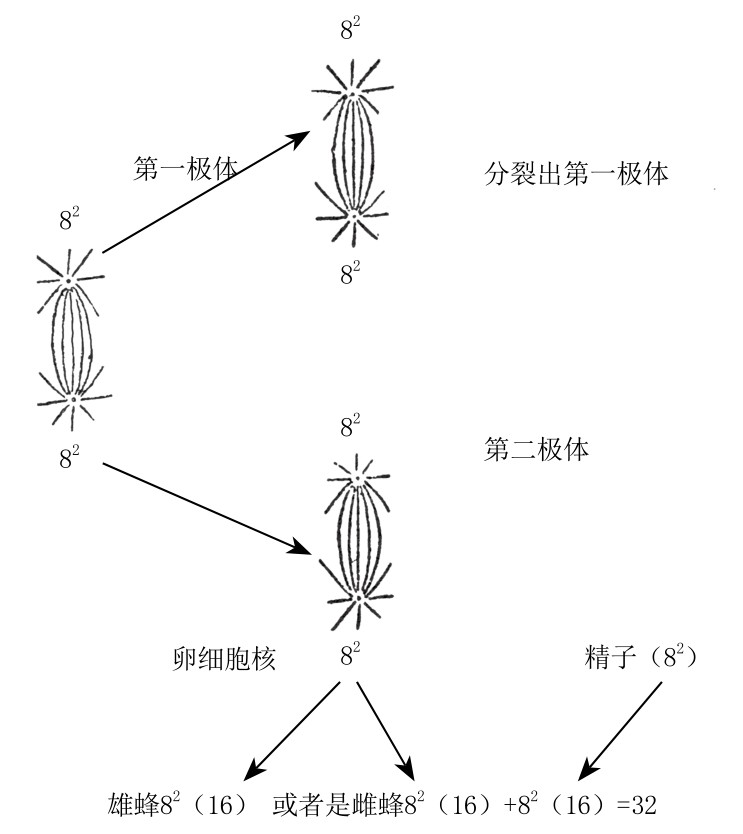
图85
蜜蜂卵细胞的两次成熟分裂。图表下端表示卵子与精子结合后,染色体一分为二,数目加倍。
鲍维里、梅林(Mehling)和纳赫茨海姆(Nachtsheim)对雌蜂和雄蜂各种组织的细胞体积与细胞核做了检测,发现:总的说来,二倍体和单倍体之间的差别一般是不恒定的。然而,无论是雄蜂还是雌蜂,在胚胎早期都出现了某种特殊现象,这使情况变得很复杂。在雌雄胚胎细胞中,一开始有两倍数目的染色体,这明显是通过染色体一分为二得到的。在雌蜂的胚胎细胞中,染色体同样一分为二,但这一分裂发生了两次,所以得到了32条染色体。这一证据似乎表明,染色体数目并没有增加,而是染色体断裂使片段变多了。如果这个解释是正确的,那么基因数目也就没有增加。雌蜂中的染色体数仍是雄蜂的两倍。对于染色体的一分为二与细胞核的大小有何关系,这一点目前尚不清楚。
不过,从雌性和雄性的胚迹看来,染色体的一分为二似乎并未发生。要是发生了,那么,这些片段在成熟期到来之前,早就重新接合了。
关于雄蜂是单倍体,或者至少其生殖细胞是单倍型的说法,细胞在成熟分裂时期的行动是最好的证据。第一次减数分裂是不成功的(如图86a,图86b),有缺陷的纺锤体将8条染色体连接起来,其中不含染色质的细胞质从中分离出来。(在第二次分裂时期)第二个纺锤体形成,而且染色体大概也通过纵裂分离开来(如图86d,图86e,图86f,图86g),两部分子染色体分别进入两极。由此,一个小细胞在较大细胞上面形成了。较大细胞变成了起作用的精子,它有着单倍数目的染色体。(https://www.daowen.com)
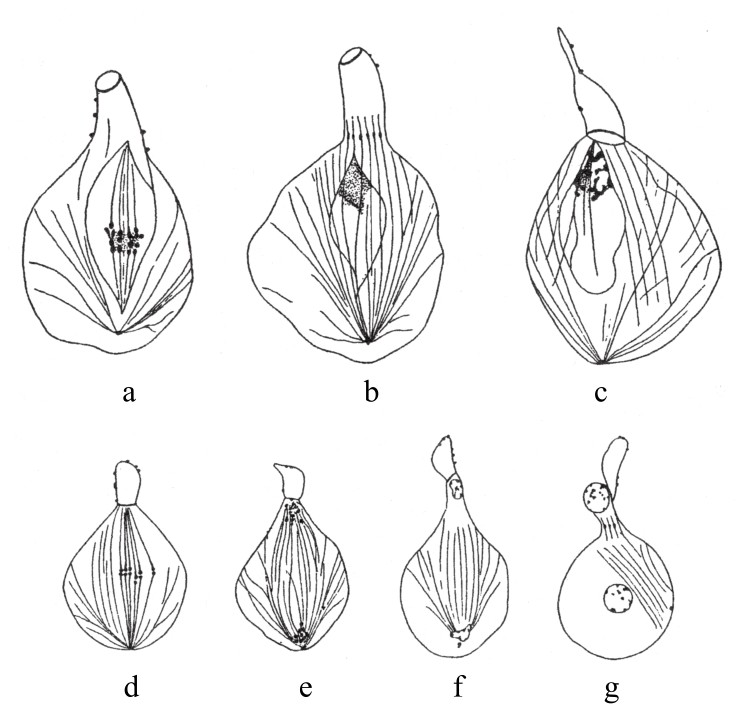
图86
雄蜂生殖细胞的两次成熟分裂。
据说,senta轮虫[6]的雄性是单倍体(如图87c),其雌性是二倍体。在食物供给差的情况下,或者说当以原生动物无色滴虫属polytoma喂养时,只会出现雌性轮虫。每只雌性轮虫都是二倍体,而且其卵细胞最开始也是二倍体。每个卵细胞都只会分出一个极体——每条染色体分裂成相同的两半。这样一来,借单性生殖所发育的卵细胞,后期会发育成雌虫,且会保留全部的染色体。当以其他食物(例如眼虫[7])喂养时,会出现新型的雌性轮虫。如果该虫在卵壳孵化时与另一只雄性结合,它只会得到能产生两个极体且保留单倍染色体数的有性卵细胞。卵细胞内的精核和卵核早就结合在一起,形成了二倍体雌性。而该雌性会重新开始一轮单性繁殖。但如果刚刚提及的这种特殊雌性并没有进行受精,那么它所产出的卵细胞会很小,且会放出两个极体,保持一半的染色体数。通过单性发育,这些卵细胞会发育成雄性单倍体。在出生几小时之后,这些雄性就会性成熟,但不再长大,且在几天之后死去。
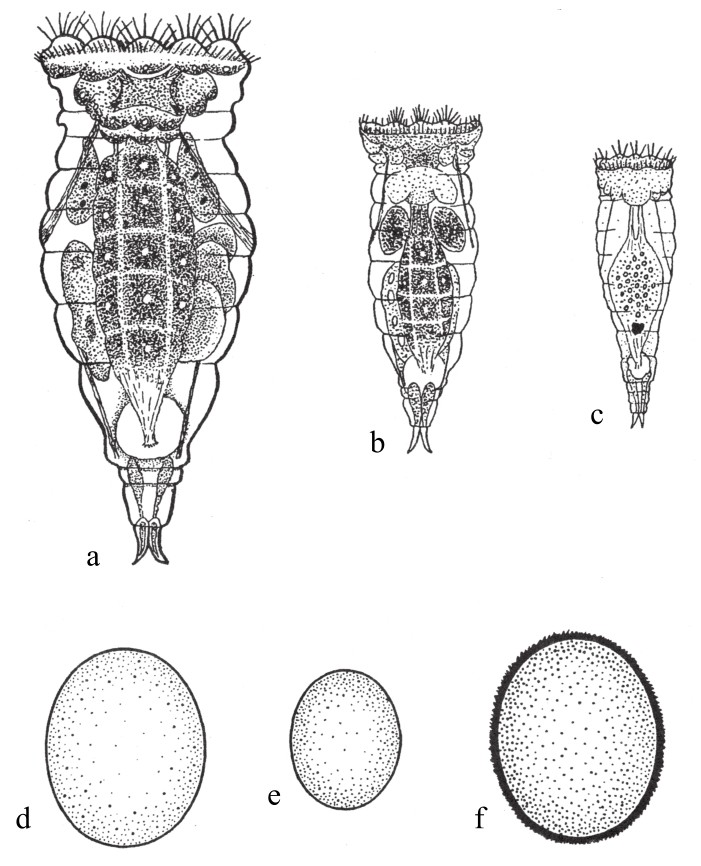
图87
图a为senta轮虫单性繁殖的雌虫;图b为其年轻雌虫;图c为其雄虫;图d为单性繁殖的卵细胞;图e为发育成雄虫的卵细胞;图f为冬季卵细胞。
施拉德(Schrader)证明了雄性vaporariorum白蝇为单倍体。莫利尔(A. W. Morrill)在美国发现,未受精的这类雌蝇只产生雄性后代。在同科的另一生物中,莫利尔和巴克(Back)也发现了相似的现象。另一方面,哈格里夫(Hargreaves)和威廉姆斯(Williams)先后报告,在英格兰,未受精的同类雌性白蝇只产生雌性后代。施拉德(1920)对美国蝇的染色体做过研究,发现雌蝇中存在22条染色体,而雄蝇中存在11条染色体。其成熟的卵细胞含有11条二价染色体,且释放出两个极体,剩下11条单染色体位于卵细胞内。如果将这个卵细胞进行受精,那么精核中的11条染色体会加入卵细胞内;如果这个卵细胞不受精,那么它会进行单性繁殖,所有胚胎细胞都会有11条染色体。在雄性生殖细胞的成熟期,看不出有减数分裂的痕迹(甚至像蜜蜂体内那样微弱的过程也没有),均等分裂也与精原细胞的分裂相同。
正如辛德尔(Hindre)的繁育实验所提示的那样,有一些证据表明,未受精的虱子卵细胞会发育成雄虱。存在一种恙虫[8]类物种bimaculatus,其未受精的卵细胞会发育成雄性恙虫,其受精的卵细胞会发育成雌性恙虫(根据几个观察者所说)。施拉德(1923)得出,这些雄虫只含3条染色体,为单倍体;这些雌虫含6条染色体,为二倍体。早期卵巢中的卵细胞有6条染色体,之后会接合成3条二价染色体。如果该卵细胞受精,精核会加入3条染色体,造就雌虫的6条染色体,于是它发育成为雌性;如果该卵细胞未受精,它会直接发育成为每个细胞都只含3条染色体的雄性。
莎尔对一种未受精的verbasci蓟马[9]雌虫做过检测,发现其只有未受精的卵细胞会发育成雄性,且这些雄性极有可能是单倍体。
苔类或藓类植株的原丝体和藓类植株世代(配子体)都是单倍体。维特斯坦用人工手段从原丝体细胞得出二倍型原丝体和二倍体藓类植株。该结果证明,这一世代和孢子体世代的差异,不是由于各世代中所含染色体数目不同而造成的,而是某种意义上的发育现象,也即这些孢子必须经历配子体状态,才能到达孢子体世代。
[1]单倍体:体细胞染色体组数等于本物种配子的染色体组数的细胞或个体。单倍体相当于原生物体细胞内染色体数目的半数体。它的基本性状虽然和原生物体相同,但一般较小,也较纤弱,植物的单倍体几乎都不能形成种子。
[2]变态:部分生物的个体在发育过程中,其形态和结构所经历的阶段性的剧烈变化,例如,一些器官退化消失,一些器官变了形态,出现新的性状。
[3]细胞分化:同一来源的细胞逐渐产生出形态结构、功能特征各不相同的细胞类群的过程,其结果是在空间上细胞产生差异,在时间上同一细胞与其从前的状态有所不同。
[4]卵裂球:受精卵发育过程中由受精卵分裂而生成的形态上尚未分化的细胞。卵裂球主要指的是从二细胞期到八细胞期之间的形态,其中每一个细胞都是胚胎干细胞,具有全能性。卵裂球之后会继续分裂形成桑葚胚。
[5]根据纳赫茨海姆的观察,未成熟蜜蜂的卵细胞中有32条染色体,但由于染色体之间两两密切接合,所以看到的只有16条染色体,这16条染色体中的每一条实际上都是二价染色体或“双染色体”。在成熟分裂以前,16条染色体又一次两两密切接合,所以他所看到的8条染色体,实际上是8条四价染色体。经过减数分裂,成熟的卵细胞各自含有8条二价染色体,实际上是16条单价染色体。精母细胞原有的16条单价染色体,并未经过减数分裂,所以成熟的精细胞各得16条单价染色体。雄蜂由卵子单性发育而成,所以只有16条单价染色体(单倍体)。蜂后与工蜂各自由受精卵发育而成,所以有32条染色体。
[6]轮虫:形体微小,长约0.04~2毫米,多数不超过0.5毫米。它们分布广,大多自由生活,也有寄生的,有个体也有群体。雌雄异体。卵生,多为孤雌生殖。
[7]眼虫:眼虫属生物的统称,在植物学中称裸藻,也称绿虫藻,是一类介于动物和植物之间的单细胞真核生物。
[8]恙虫:又称恙螨、沙虱,能传染恙虫病。在动物分类上,恙螨属节肢动物门蜘蛛纲。全世界已知的恙螨有3 000多种,中国有记载的达350余种。
[9]蓟马:昆虫纲缨翅目的统称。幼虫呈白色、黄色或橘色,成虫呈黄色、棕色或黑色;取食植物汁液或真菌;体微小,体长0.5~2mm。