突变性状的起源
第五章 突变性状的起源
进化必须使基因发生改变才能得以进行。然而,我们并不是说进化性的改变和由突变引起的改变是一样的。极有可能的是,野生型基因有其不同的起源。事实上,我们已经接受了这一观点,有时还很热衷于主张它。因此,找出是否存在可以支持这一观点的证据,意义重大。
现代遗传研究与新性状的起源,有着十分密切的联系。事实上,只有当数对用于比较的性状可被追踪时,孟德尔式的遗传研究才有可能继续进行下去[1]。孟德尔在他所使用的商品豌豆中找到了高茎矮茎、黄皮绿皮、圆粒皱粒这三对性状。虽然之后的研究也大量使用了这些材料,但一些最好的研究材料,却是其谱系中起源较为确定的新型性状。
这些新性状大多数是突然出现的,且具备完整的遗传能力,和其原型性状一样稳定。例如,在培养过程中,会突然出现白眼性状的雄果蝇,若让它和普通的红眼雌果蝇交配,所得子一代全为红眼果蝇(如图38)。子一代杂交所得子二代中又会出现红眼和白眼的个体,且所有白眼的个体都是雄蝇。
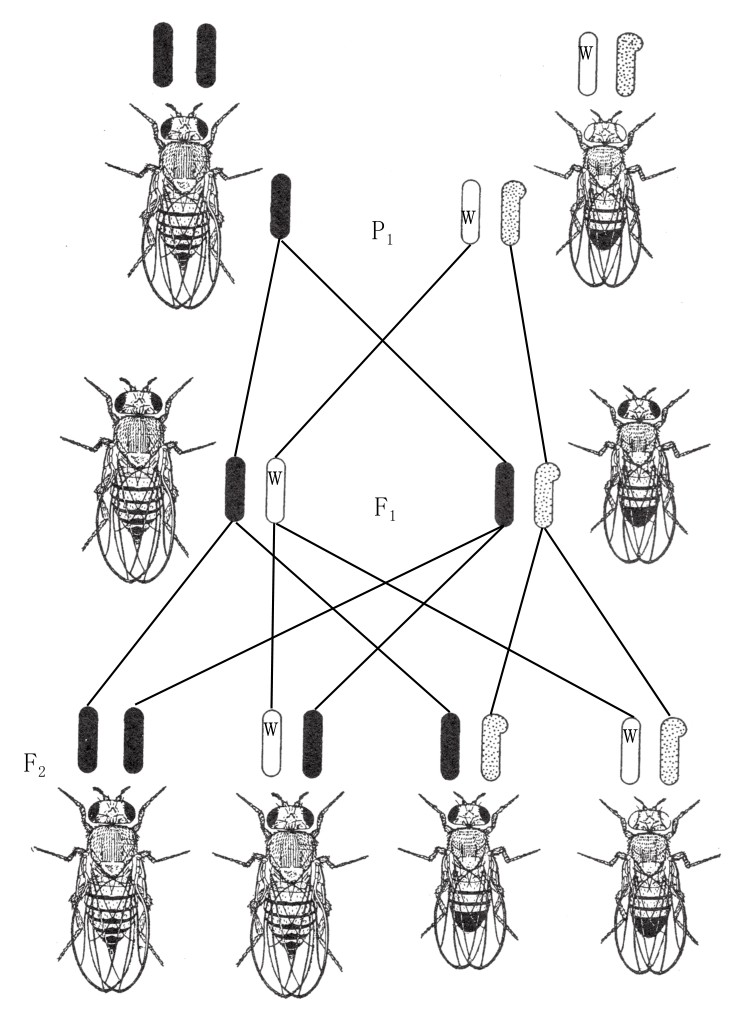
图38
黑腹果蝇的白眼性连锁遗传。一只白眼雄蝇与一只红眼雌蝇交配。携带红眼基因的X染色体用黑棒表示;携带白眼基因的X染色体用白棒表示;Y染色体用斑点棒表示;在染色体上,白色隐性基因用字母w标出。
接下来,将这些子二代白眼雄蝇和同代(子二代)的红眼雌蝇交配,所得后代与子二代中白眼和红眼数量相同,且后代白眼果蝇中,既有雄蝇也有雌蝇。如果这些白眼果蝇自交,所得后代均为纯合子白眼果蝇。
我们根据孟德尔第一定律对这一实验作出解释,即假设在果蝇的生殖质内存在产生红眼和产生白眼的两种要素(或基因),它们是一组相对要素,会在杂合子的卵细胞或者精细胞成熟时分离开来。
值得注意的是,我们所观察到的这一点,并不表明白眼基因单独产生白眼性状。它只表明如果原始物质发生了部分改变,会导致整个物质催生出一种不同的最终产物,即我们所称的新性状。实际上,这一改变不仅影响了眼睛的性状,也影响了躯体的其余部位。红眼果蝇的精巢膜原本是带了点绿色的,但白眼果蝇的精巢膜却呈现为无色;白眼果蝇与同类红眼果蝇相比,行动较为迟缓,而且白眼果蝇的寿命更短。这可能是因为其生殖质内发生了一些变化,这些变化使身体的很多部位受到了影响。
自然界中的醋栗蛾,有些罕见的个体呈现浅色或白色,一般情况下,它们都是雌性。浅色的突变型雌蛾和黑色的野生型雄蛾交配(如图39),所得子一代的性状与黑色野生型的相同。子一代自交所得子二代中有新旧两种类型的蛾,且浅色蛾与黑色蛾比例为1:3。子二代个体中浅色蛾全为雌性。如果将子二代中的浅色雌蛾与同代雄蛾交配,会得出雄性和雌性同样多的浅色醋栗蛾,也会得出雄性和雌性同样多的黑色醋栗蛾。我们可以从这一代开始培养浅色醋栗蛾。
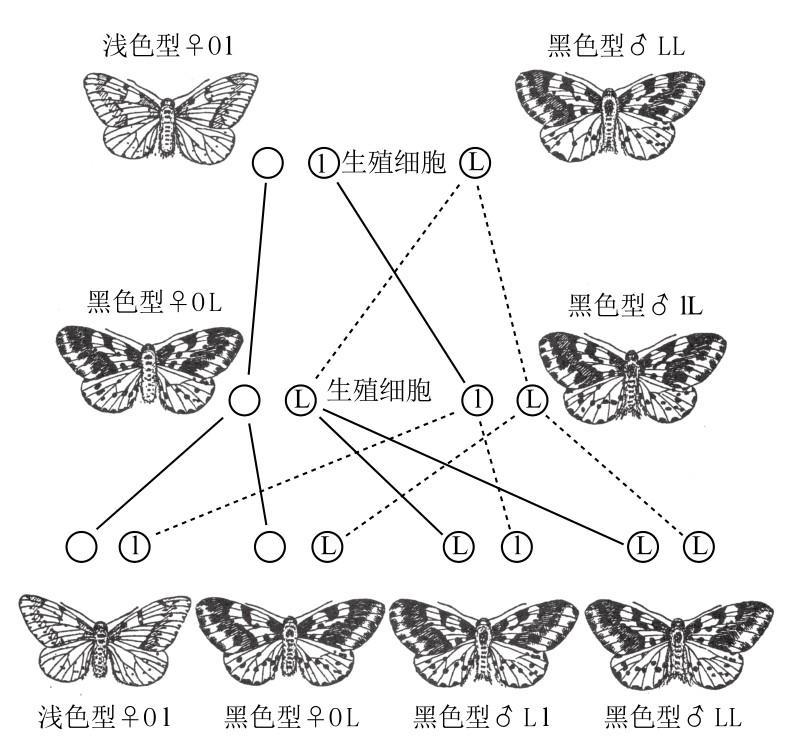
图39
浅色型醋栗蛾和黑色型醋栗蛾的杂交。携带黑色基因的性染色体用带L的圆圈表示;携带浅色基因的性染色体用带l的圆圈表示;雌性蛾所独有的W染色体用无字母的圆圈表示。
以上两种突变性状相对于野生型性状表现为隐性性状,但也有另外的突变性状相对于野生型性状表现为显性性状,譬如,果蝇Lobe2眼的性状特征在于眼的特殊形状和大小(如图40)。这一性状,起初只是出现在某个果蝇身上。这种性状的果蝇所孵出的后代中,有半数有此Lobe2眼。在突变型的父方或是母方体内,有一条第二染色体,一定在某处发生了基因突变。在受精时,含此突变基因的生殖细胞与一个正常的生殖细胞结合,第一个突变个体由此而来。因此,第一个Lobe2眼的果蝇是杂种或杂合子。并且,就像以上所陈述的一样,当此杂合子与正常的个体交配时,所得后代会有Lobe2眼和正常眼两种果蝇,其数量各占后代的一半。让杂合子Lobe2眼果蝇进行自交,会得到纯种的Lobe2眼果蝇,但通常情况下,其眼睛更小,且有可能缺失一只眼睛甚至没有眼睛。
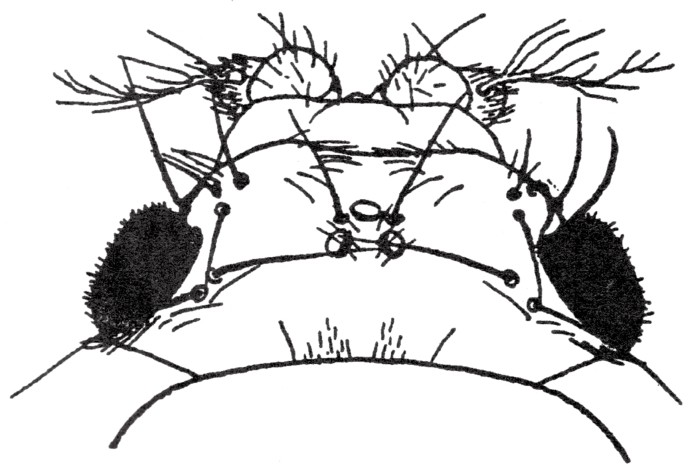
图40
黑腹果蝇的突变性状Lobe2,眼小而突出。
有一个很奇怪的现象,那就是当突变显性基因以纯合子的形式出现时往往是致命的。因此,当果蝇带有纯合子的卷翅(如图41)突变基因时,它们几乎都会死亡,只有极少的个体能存活下来。老鼠的黄毛突变显性基因以纯合子的形式出现时,是致命的;老鼠的黑眼和白毛突变基因也是如此。在所有携带突变显性基因的物种中,都不可能产生纯种的个体(除非出现另外一种致死基因与这一突变显性基因相抵)。在它们所产生的后代个体中,每一代中都有一半像它们自己,而另一半表现为另一正常型(携带正常的等位基因)。
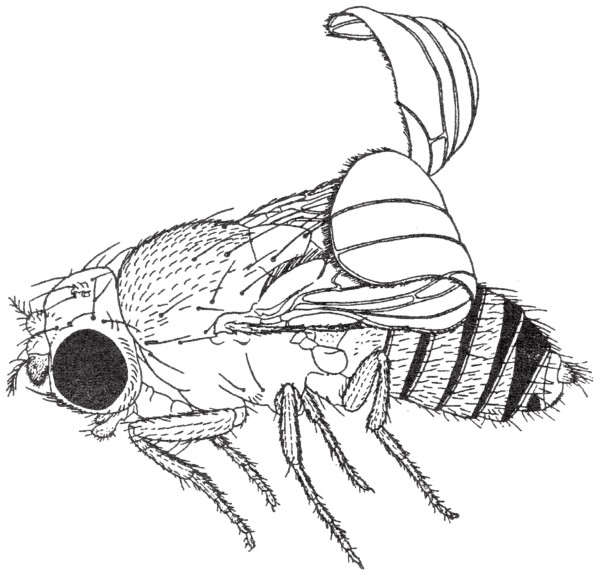
图41
黑腹果蝇的突变卷翅性状:翅尾往上翻卷,且两翅稍微分离。
众所周知,人类的短指症[2]是一个突出的显性性状。毋庸置疑,短指自身作为一个显性的突变性状,会在有着短指性状的某些家族中持续遗传。
所有果蝇的原种,都是以突变体的形式出现的。在以上我们所列举的例子中,(后来以原种的形式固定下来的)突变体一开始出现都是一个个体。然而,在别的例子中,存在多个个体同时表现出新的突变性状。这些突变准是在种系[3]早期就发生了,所以才会出现几个卵细胞和几个精细胞携带这些突变要素的情况。
有时,一对果蝇会有四分之一的子代都是突变体。而且有证据显示,这种突变早已在某一祖先体内就出现了。但由于这些突变体都是隐性的,所以,如若不是含有这一突变基因的两个个体交配,这种突变性状便不会显现出来。因此,在它们(自交)的子代中,预计会有四分之一的个体显现出这种隐性性状。
近亲繁殖的种群比远亲繁殖的种群更易产生这种结果。因为如果是远亲繁殖,在两个个体偶然会合之前,隐性基因可能已分布到大量的其他个体中去了。
人类存在一些由突变引起的缺陷性状,且其重现的次数要比预期的更高,这极有可能是因为在人体的生殖质中隐含着很多隐性基因。如果对人类的遗传谱系予以追踪,会发现其亲戚或祖先也有着同样的突变性状。或许,人类的白化病[4]是证明这一观点的最好例子。在很多白化病案例中,白化病多来源于父母双方都携带着致病的隐性基因,但新的白化基因也有可能是由突变产生的。即便如此,直到它遇到另外一个相同的隐性基因之前,这个(隐性)突变的白化基因的性状是不可能表现出来的。
人类驯化的动植物所展现出的众多性状,都会像起源已确定为突变的那些性状一样,一代一代固定地遗传下去。毋庸置疑,很多性状源于突变,尤其是那些由近亲繁殖而来的驯化类型。
当然,我们并不能由此断定,只有驯化的品种才会产生突变,因为事实并不是这样。有大量证据表明,在自然界同样也有此类突变体。但由于大多数的突变体比野生型更虚弱,且适应性更差,以致这些突变体在被认出之前就已经消亡了。相反,在人工培育过程中,这些个体受到了保护,虚弱的个体也有机会存活下来。加之驯化动植物,尤其是那些供遗传研究而培育的动植物,都经过了仔细的检查,我们对其(表现出的性状)非常熟悉,所以能觉察出新类型。
一项对黑腹果蝇原种突变的研究,揭露了一个奇怪的意外事实:在果蝇的一对基因中,仅有一个基因发生突变,而不是一对基因中的两个同时发生突变。究竟是什么类型的环境导致一个细胞中的某个基因改变,而另一个等位基因却没有发生任何改变,这是很难想象的。因此,我们推断,引起改变的原因似乎是内在的,而不是外在的。稍后,对这一问题,我们将做进一步探讨。
另一事实也在研究突变作用的过程中引起了我们的关注,即相同的突变可能一次又一次地重现。表1是果蝇突变重现的列表。相同突变的重现表明我们所研究的是独特而有序的过程。突变的重复出现让我们想起了高尔登(Galton)著名的多面体比喻。多面体的每一个变化都与基因的某个新的稳定位置相对应(这里或者用在化学意义上)。
表1 重现的突变和等位基因系
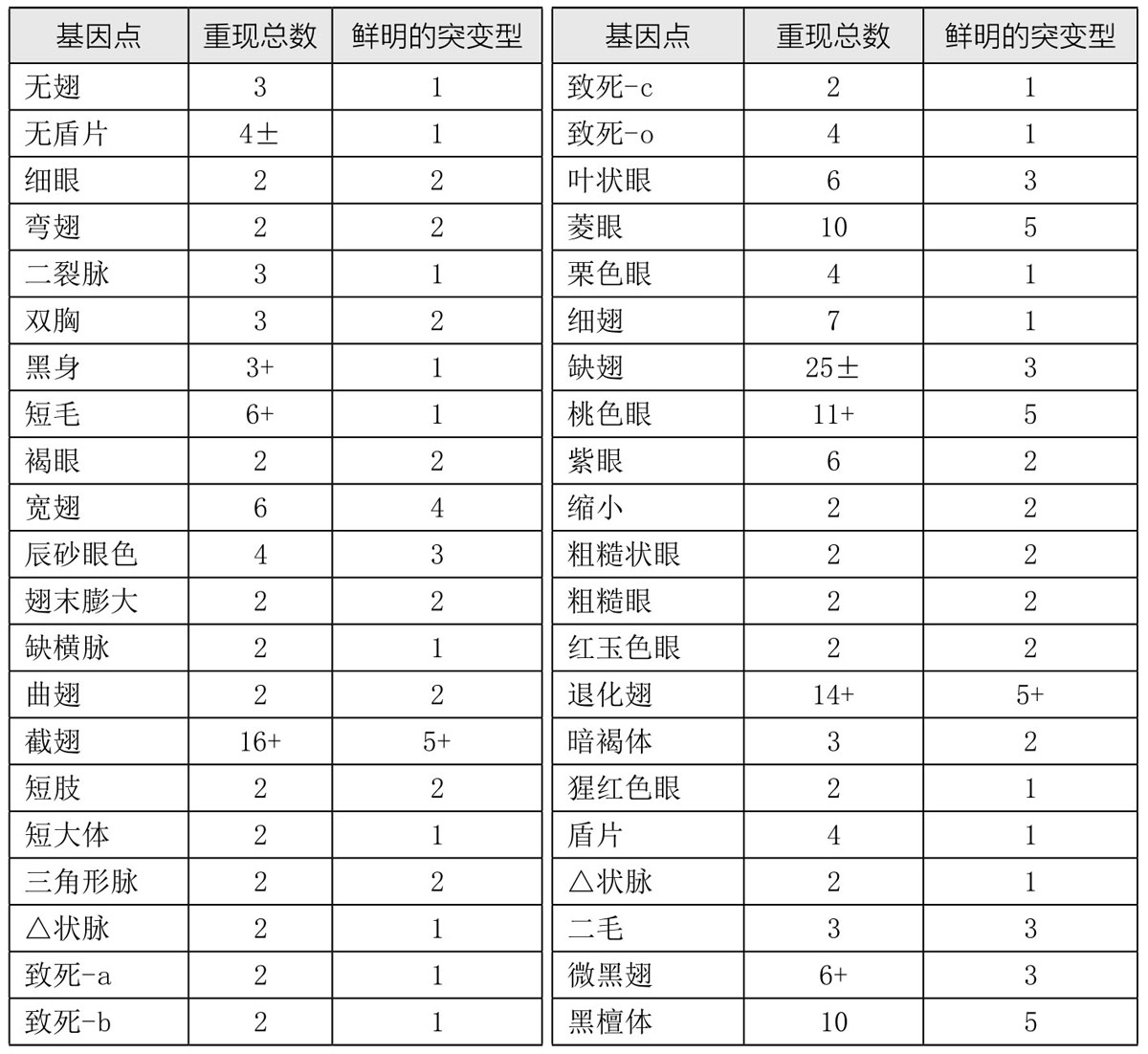
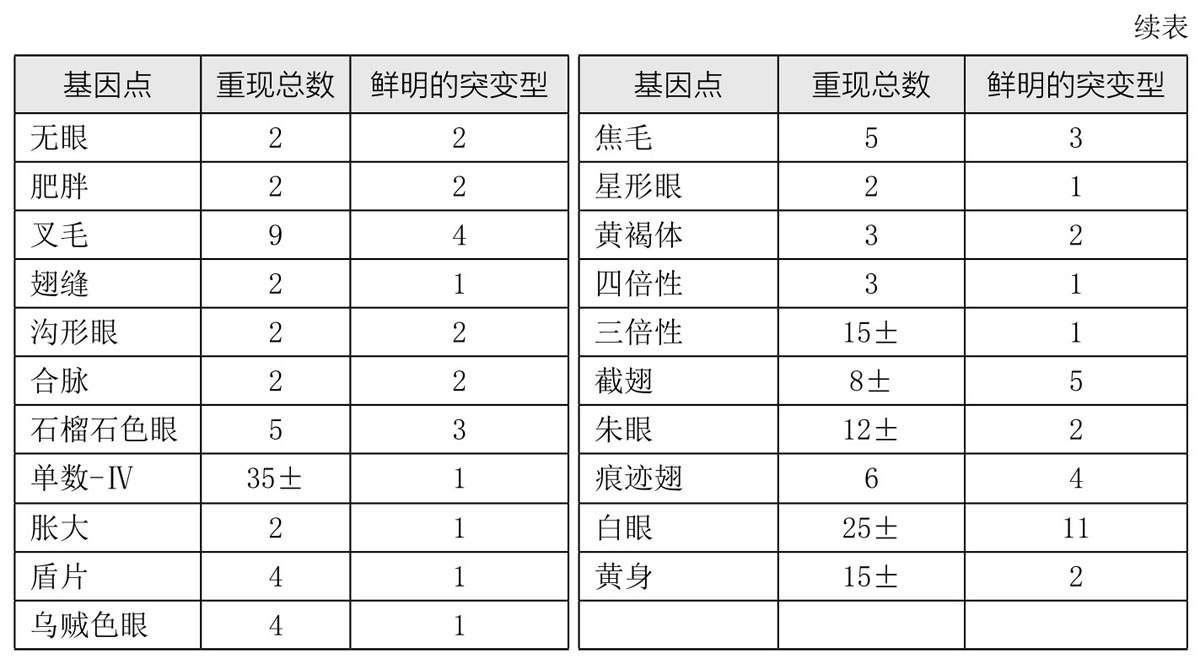
我们最常引用或用作遗传学研究资料的突变性状,通常都是那些相当激烈的改变或畸形。这就给人留下了这样一个印象,即突变型与原始型是有很大差距的。当达尔文谈及飞跃[5]时,他并没有把这些飞跃当作生物进化的证据,因为他说,如此大的改变有可能使得躯体的部分不太能适应已经调和了的环境。一方面,当激烈的变化使基因突变或导致个体畸形时,我们会充分意识到达尔文的这一论点是正确的;另一方面我们也意识到,小的改变和大的变化一样,都是突变的特征。事实已多次证明,一部分稍大或稍小的轻微改变,都可能是源于胚胎质内某些基因的改变。既然只有源于基因的差异才能遗传,那么,结论似乎是:进化必须通过使基因发生改变才能得以进行。然而,我们并不是说进化性的改变和由突变引起的改变是一样的。极有可能的是,野生型基因有其不同的起源。事实上,我们已经接受了这一观点,有时还很热衷于主张它。因此,找出是否存在可以支持这一观点的证据,意义重大。表面看来,在德弗里斯(de Vries,1848—1935)[6]的著名突变论[7]的早期论述中,似乎就已暗示了新基因的产生。(https://www.daowen.com)
突变论一开始就提出:“有机体的性质是由单元组成的,而这些单元是截然不同的。它们会结合成群,且在近缘种(相近物种)中,相同的单元和单元群组会重现。但在动植物外形上是看不见这样的过渡阶段的,因为发生在单元和单元之间的转换,就像发生在化学中分子和分子之间的变化一样是看不到的。”
“物种间并没有连续性的联系,新物种的出现源于突然的变化或层级。当新的单元加到原有单元组时,就会形成一个层级,于是新型成为独立种,并从原有物种中分离出去。这里的新物种便是那个‘突然变化’。这一新物种的出现,看不出有任何准备,也没有过渡。”
以上观点似乎是说,有一个突变就会产生一个初级的新物种,而突变之所以会产生,是因为一个新要素即新基因的突然出现。还有另外一种说法:我们见证了突变时诞生的新基因,或者至少见证了新基因的活动,世界上具有活性的基因的数目,因此增加了一个。
德弗里斯在其《突变论》的最后几章和他后来关于“物种和变体”的演讲中,进一步阐释了他对突变的理解。他承认两种作用的存在:其一,新要素(即基因)的增添会带来新物种;其二,原有的要素(基因)会失去活性。目前,我们对第二个作用较感兴趣,因为除了表达方式不同以外,第二种作用实质上是在陈述这样一个观点:今天我们培育的新型,是源于一个原有基因的缺失。事实上,德弗里斯自己将所有这些常见的缺失突变——不管其是显性的还是隐性的,都列入了这一作用,但他自己默认这些突变都是隐性的,毕竟它们的基因都失去了活性。他认为出现孟德尔式结果是因为有好几对相对基因——具有活性的基因及其无活性的等位基因,所以都属于第二种作用。每对基因中的等位基因彼此分离,于是便有了孟德尔式遗传中所特有的两种配子。
德弗里斯还提到,这样一个过程代表了进化向“后”迈了一步。这不是进步,而是退步,且产生了一个“退化变种”。就像我们提到的那样,这种解释和目前所主张的物种突变是由于基因缺失的说法类似——原则上,两种说法是一样的。
因此,检测那些促使德弗里斯推进其突变假设的证据是有意义的。
在荷兰首都阿姆斯特丹的一片荒地上,德弗里斯发现了一丛拉马克待霄草(Oenothera lamarckiana)[8](如图42)。其中有几株较为特殊,它们与普通的植株稍有不同。他将这些特殊植株移植到自己的花圃,发现大多数能继续繁育特殊植株。同样,他也种植了一些祖代植株,或普通型拉马克待霄草。在这些祖代的后代里,每一代也产生了少量相同的新型植株。当时,总共有九种这样的植株被鉴别出来,且都是新的突变型。

图42
左图是普通型拉马克待霄草,右图是巨型待霄草。
现已证实,其中一种突变型是因为染色体数量加倍,这种突变体被称为巨型(gigas)待霄草[9](如图42)。有一种突变型是三倍体,被称为半巨型(semi-gigas)待霄草。有几种突变型是因为出现了额外的染色体,被称为lata型待霄草和semilata型待霄草。至少有一种brevistylis待霄草和果蝇的隐性突变一样,两者都属于基点突变。而对于brevistylis待霄草和隐性突变的残余待霄草,才是德弗里斯所能用于研究引用的[10]。现如今,很有可能这种残余类型(隐性突变体)和果蝇突变类型是相似的,但它们在近几代中都会重现,这一点又与果蝇以及其他动植物的突变情况完全不同。有一个合理的解释或许可以说明这一现象,即这些隐性突变基因与存在的致死基因是紧密连锁的。只有当这些隐性基因通过交换作用,从附近的致死基因中分离出来时,这一性状才能表现出来。在果蝇中,有可能存在含隐性基因且与待霄草相似的平衡致死纯种。所以,只有当交换发生时,这些隐性基因才会重现,且其重现频率决定于致死基因和隐性基因之间的距离。
已有发现表明,野生待霄草的其他物种也会表现出和拉马克待霄草一样的行为,因此,我们可以看出,拉马克待霄草的遗传属性与其杂交种的起源无关(像我们有时推想的那样),而主要是由隐性基因与致死基因连锁导致的。突变型的出现,并不代表隐性基因发生突变的过程,而代表隐性基因脱离致死基因的连锁而得到解放的过程[11]。
这样看来,拉马克待霄草的突变过程,同我们所熟悉的其他动植物的突变过程,似乎没有本质区别。换句话说,除了因为致死基因和一些隐性突变基因之间有连锁关系外,发生在拉马克待霄草身上的突变过程,同发生在动植物身上的突变过程,在解释上不会有什么本质区别。
综上所述,我认为,即使出现一个新型或者是进步型待霄草品种,也没有必要假设增加了一个新基因。德弗里斯所提出的进步型,或许就是在正常的常染色体中多出来一条额外的染色体。这一问题将在第十二章继续讨论,目前我们需要指出的是:很少有证据可以表明,新物种往往是通过增加染色体的途径而产生的。
[1]研究所考虑的遗传特征必须是等位基因对的表达结果,其中一个基因相对另一个基因呈显性。分离定律指出,体细胞中的基因是成对出现的,而每一个配子只含有每对基因中的一个基因。自由组合定律指出,如果具有一对以上基因,那么每一对基因的分离都与其他各对基因的分离无关。虽然这些定律组成了现代遗传学的基础,但它们并不总是正确的。它们只能适用于那些没有被其他位点上的基因的影响修饰过的基因对或并不在同一个染色体上的基因对。同一染色体上的基因对的分离受交换程度的影响。
[2]短指症:又名短趾症,是一种常染色体显性遗传病。
[3]种系:假定有共同起源且关系密切的一小群物种。一般不患病。
[4]白化病:由于酪氨酸酶缺乏或功能减退引起的一种皮肤及附属器官黑色素缺乏或合成障碍所导致的遗传性白斑病。患者视网膜无色素,虹膜和瞳孔呈现淡粉色,怕光,皮肤、眉毛、头发及其他体毛都呈白色或黄白色。白化病属于家族遗传性疾病,为常染色体隐性遗传,常发生于近亲结婚的人群中。白化病患者双亲均携带白化基因,本身不发病。如果夫妇双方同时将所携带的致病基因传给子女,子女就会患病。眼白化病为X连锁隐性遗传,当母亲所携带的白化基因传给儿子时儿子才会患病,其女儿一般不患病。
[5]飞跃:一种激烈的突变。
[6]德弗里斯:荷兰植物学家和遗传学家。他早年研究植物生理学,在渗透压方面成果卓著。1873年他所发表的两篇关于攀援植物运动机制的笔录,受到达尔文赏识(见达尔文著:《攀援植物的运动和习性》)。他后来转向遗传学研究,是孟德尔定律的三个重新发现者之一。
[7]1901年,德弗里斯首次提出生物进化起因于“突变论”的观点,他认为新物种是通过一系列急剧变动、突进或跳跃式变异即突变而出现的。后来的遗传学研究表明,德弗里斯所说的突变主要指的是染色体畸变。
[8]又称拉马克月见草。两年生草本,茎多分枝,高达1米,具白长毛,毛基部带红色突起。
[9]即四倍体待霄草植株。
[10]德弗里斯和施通普斯(Stomps)都认为,巨型待霄草的一些特征是源于除染色体数量外的其他因素。
[11]沙尔(A. F. Shull)已经在致死基因连锁假设上解释了大量拉马克待霄草的隐性基因类型。埃默森(S. H. Emerson)最近指出,沙尔所发表的证据虽然不能完全令人信服,但也可能是合理的。在最近的出版物中,德弗里斯自己似乎也没有对采用致死基因连锁假说来解释多次重复的隐性突变基因这一点提出反对。他还将隐性突变基因置于“中央染色体”内。