基因的稳定性
第十八章 基因的稳定性
如今,我们把选择(不论是自然选择还是人为选择)所引起的这种变化,最多视作只在原有基因组合能够影响的变化范围内可引起的变化。换言之,选择不能使一个组群(物种)超过这一组群原有的极端类型。严格的选择能使种群达到一个点,在这个点上几乎所有个体都接近原群表现的极端类型,但超越了这个点,选择就不起作用了。现在看来,只有一个基因内部产生了新的突变,或者是一群原有基因内部出现了集体改变,才有可能引发永久性的变异,但这一变异有可能是进一步,也有可能是退一步。
至今,我们所谈到的种种,都暗示着基因在遗传上是一个稳定的要素。至于基因的稳定性是基于化学分子的哪一种稳定,或者只是在一个恒定的标准数值附近定量地上下波动,却是理论上或根本上的一则重要问题。
既然不能通过物理方法或化学方法直接研究基因,那么,我们关于基因稳定性的结论,就只能从其所产生的效应来加以推导。
孟德尔遗传理论假设基因是稳定的,它假设父方和母方赋予杂合子基因,在新环境中基因会在杂合子体内完整地保留下来。现在,让我们借助几个例子来回忆一下这一结论的实质性证据。
安达卢西亚鸡有白色、黑色和蓝色三种类型。如果让白鸡和黑鸡交配,子代为蓝灰鸡或蓝鸡。让两只蓝灰鸡交配,所得子代为黑鸡、蓝鸡和白鸡,且比例是1:2:1。在杂合子蓝鸡中,白色基因和黑色基因是分离开来的。一半成熟的生殖细胞会携带黑色基因,一半成熟的生殖细胞会白色基因。这些精细胞和卵细胞随机结合,据观察,孙代不同体色的比例是1:2:1。
为了验证杂合子生殖细胞中存在两种基因的推测是否正确,我们做了如下测试:用杂合子蓝鸡回交纯种白鸡,所得子代会有一半蓝鸡和一半白鸡;用杂合子蓝鸡回交纯种黑鸡,所得子代会有一半黑鸡和一半蓝鸡。可见,两次结果都与这一假设相符,即杂合子蓝鸡的基因是纯的,其中一半为黑色基因,另一半为白色基因。这两种基因出现于同一个细胞内,相互之间没有污染。
在刚才给定的例子中,所得杂合子既不像父方也不像母方,从某种意义上来说,是介于父方和母方之间的中间品种。然而在下面的这个例子里,杂合子和祖代中的一方是没有区别的。如果黑豚鼠[1]和白豚鼠交配,所得后代(子一代)全为黑豚鼠。让所得黑豚鼠自交,所得后代(子二代)豚鼠的黑白比例为3:1。子二代的白豚鼠又能像祖代的白豚鼠一样,繁育出白豚鼠。从这个例子中也可以看出,虽然在杂合子中,白色基因和黑色基因共处了一段时间,但白色基因并未因此而被污染。
下面这一案例,祖代的双方极其相似。虽然所得杂合子表现出某种程度的中间型,但变动很大,其性状慢慢往祖代两方的性状靠拢,最后与祖代两方重合。在这种类型里,只有一对基因存在差异。
如果让黑檀色果蝇与乌黑色果蝇交配,其子一代正如上文所述,呈现出不同的中间体色。如果让子一代的杂合子自交,所得子二代的体色由浅到深,实际上呈现为一个连续的系列。然而,也有一些测试体色等级的办法。当子代的杂交完成后,我们发现其子二代的个体有纯黑檀色果蝇,有杂合子果蝇,还有纯乌黑色果蝇,且比例为1:2:1。在此,我们又一次得到基因稳定而未相互污染的证据。这一系列由浅到深的体色,仅仅是由于不同性状的叠加而形成的。
所有的这些例子简单明了,因为我们所涉及的每一个例子都只涉及单组基因所表现出来的差异。这些例子都为证明基因的稳定性原则服务。
然而,现实情况中往往不会一直这么简单。很多类型,在几组基因上都有差异,每组基因对同一个性状都有影响。因此,在它们的杂交后代里,无法得到简单的比例。例如,用短穗玉米与长穗玉米杂交,子一代会得到中穗玉米。将中穗玉米自交,子二代中会有各种穗长不一的玉米。有一些玉米穗,同原来的短穗族一样短,有一些同原来的长穗族一样长。在长穗和短穗之间,有一系列长短不一的中穗玉米。研究者检测子二代的个体,发现有几组基因对穗长存在影响。
还有人类身高的例子。某人的身体高,可能是因为腿长,可能是因为躯干长,又或者是两部分都长。有些基因可能会影响所有部分,而另一些基因可能对某个部位有着相比于其他部位更大的影响。结果便使得遗传情况复杂化,且至今未获解决。此外,环境在某种程度上也可能影响最终的结果。
这些都是多对基因参与的例子,遗传学学者还在努力探究每一次杂交中到底有多少基因参与性状的表达。单单因为涉及了几个基因或数个基因,这一结果就已相当复杂了。
在孟德尔的发现被披露前,正是这一类变异性,为自然选择理论提供了证据。这一问题,我们还是留待以后再行探讨吧。首先我们需要探讨一下的是,在认知选择学说中的一些限制上,由约翰森(Johannsen)[2]在1909年的杰出研究中所取得的巨大进步。
约翰森用一种园艺植株,即公主菜豆来做实验。这种公主菜豆,严格以自花授粉的方式来繁育后代。因此,长期的自交使所有个体都变成了纯合子。这就是说,一对基因中的两个因子都是相同的。因此,公主菜豆适合用于进行精密的实验,借以判定菜豆间个体差异是否受选择的影响。如果选择改变个体的性状,那么,在这种情况下,性状的改变肯定是通过基因自身的改变而形成的。
不同植株所产出的豆粒在大小上有一定的差异。当将其按照大小顺序排列时,其数量会呈现为正态分布曲线。不管从每一代中选出来繁育后代的是大豆子还是小豆子,任一植株以及它的所有后代,在豆粒的大小上,都有着相同的分布曲线(如图150)。其后代总会得到(大小分布规律)相同的豆群。
约翰森在他检测过的菜豆中发现了九族菜豆。他认为,对这种菜豆的研究结果足以证明,给定植株之所以得到不同大小的菜豆,很大程度上是由其所处的环境因素所导致的。当选择开始时,只需要选用有着同组基因的材料,便可以证实这一论点。这说明,选择对基因自身的改变没有影响。
如果一开始选择的有性生殖动物或植物不是纯合子,那么,直接结果就不同了。为了证明这一点,研究者做了大量实验,例如居埃诺(Cuènot)对斑毛鼠的实验,或麦克道尔(Mac Dowell)对家兔耳长的实验,又或者伊斯特(East)和哈耶斯(Hayes)对玉米所做的实验。从中任选一个实验,都可以作为例子,来说明在选择下所发生的变化。在此只详述一例。
卡斯尔(Castle)研究了选择作用对一族披巾鼠(也称条纹鼠)毛色的影响(图151)。他开始用市场售卖的披巾鼠的子代做实验,他选择了两只披巾鼠,分别代表两方,一方是宽条纹的,另一方是窄条纹的。他将这两只披巾鼠分开饲养。在经历了几个世代的繁育之后,这两群披巾鼠出现了些许不同:宽条纹的一方,后背条纹越来越宽,其平均宽度甚至超过了原始披巾鼠的条纹宽度;窄条纹的一方,后背条纹的平均宽度,也比原始披巾鼠的条纹更窄了。选择作用确实以某种方式改变了条纹的宽度。到目前为止,这些实验结果还不能证明:这一改变不是由于选择作用将两组决定后背条纹宽度的基因分开而引起的。然而,卡斯尔认为,他研究的是单个基因的效果,因为当这些披巾鼠与全黑鼠或是全褐鼠杂交时,所得子一代会是杂合子。再用子一代的杂合子自交,所得子二代便会得出全黑鼠(或全褐鼠)与披巾鼠两型,比例为3:1。事实上,这一孟德尔式的比例表明,披巾鼠毛上的有色条纹起源于一个隐性基因,但该结果未能证明这一基因所表达的效果有可能受到其他基因的影响,而这些其他基因决定着后背条纹的宽度。这才是真正存在争议的问题。

图151
四个类型的披巾鼠。
之后,卡斯尔完成了由弗里格特(Wright)设计的一个实验,事实证明:这些结果的出现,是条纹宽度的修饰基因被隔离开来所造成的。实验方法如下:让选择出来的披巾鼠种群(宽条纹和窄条纹的披巾鼠)与野生型披巾鼠回交(全黑或全灰的披巾鼠),挑选得出第二代(子一代)条纹披巾鼠。再将所得条纹披巾鼠与野生型披巾鼠回交,如此反复,直至回交两三代之后,我们发现,所挑选的条纹披巾鼠开始往回发育,也就是说回到了最开始的状态。我们所挑选出的窄条纹披巾鼠,慢慢往宽条纹披巾鼠的方向变化,而宽条纹披巾鼠慢慢往窄条纹披巾鼠的方向变化。换句话说,精选的两族披巾鼠彼此越来越相似,越来越像它们最初的那一族披巾鼠。
这一实验结果完全符合以下观点,即存在于野生型披巾鼠中的修饰基因会影响条纹鼠已有的条纹宽度。换句话说,原来的选择作用,通过分离变宽基因和变窄基因,从而改变披巾鼠条纹的宽度。
曾有一段时间,卡斯尔甚至自认为披巾鼠的实验结果重建了达尔文自然进化论的观点,即选择会使得遗传物质往所选方向发生改变。如果这真的是达尔文的本意,那么这一观点对变异的解释,看似可以壮大如下理论:进化是借自然选择而进行的。卡斯尔在1915年说:“我们目前所拥有的所有证据表明,外界的修饰因子不能说明我们在披巾鼠上所观察到的变化,而披巾鼠的变化是一个明确的孟德尔式单元。由此,我们被迫得出以下结论,即这一单元自身是在反复选择下,向着自然选择的方向改变的;有时这一改变很突然,就像‘突变族’一样,而突变族自身是一种高度稳定的正向突变[3];但更常见的改变是慢慢改变,就像在正负两个选择序列中不断发生的那样。”
一年后,他说:“很多遗传学的学者目前都将单元性状看作一成不变的……这一问题,我已研究多年,在这一点上得出了一个大致的结论,即这些单元性状是可改变的,也是可重组的。很多孟德尔学派的人并不这么认为,在我看来,那是因为他们对这一问题的研究不够深入。事实很明显,单元性状会发生定量变异。……选择作为进化的动力,需被重新放到达尔文预言过的重要位置;选择作为动力,能持续推动种族发生进步性的改变。”
细看达尔文的著作之后,我们会发现达尔文进化论并非像卡斯尔所说,选择会决定或是影响未来的变异方向。除非我们引用达尔文关于获得性遗传理论的另一项假说(即泛生论),否则是找不到一点证据可以佐证卡斯尔的观点的。
达尔文坚信拉马克学说[4]。无论何时他的理论遇到困难,他都会立刻用拉马克学说去支撑其观点。因此,任何人只要愿意(尽管达尔文并未将两种观点混为一谈,卡斯尔也没有),都可以极具逻辑性地指出,无论何时,一个更有优势的类型被选择,其生殖细胞都会接触到其体细胞所产生的泛子,且受到泛子的影响,预计会往所选性状的方向改变。因此,每一次新进展都源于一个新的基础。如果突变散乱地发生,那么势必会出现一个新的模式,可以超越先前的界限,往上一次革新所发生的那个方向上延展。换句话说,自然选择会在每种选择发生的方向上往更远的方向发展。
虽然人们认为,每当达尔文发现自然选择学说有不能解释的情景时,他在原则上便会借拉马克学说来为新的进展提供支持,但是,就像我所说的那样,达尔文从未借拉马克学说来支持他的自然选择学说。
如今,我们把选择(不论是自然选择还是人为选择)所引起的这种变化,最多视作只在原有基因组合能够影响的变化范围内可引起的变化。换言之,选择不能使一个组群(物种)超过这一组群原有的极端类型。严格的选择能使种群达到一个点,在这个点上几乎所有个体都接近原群表现的极端类型,但超越了这个点,选择就不起作用了。现在看来,只有一个基因内部产生了新的突变,或者是一群原有基因内部出现了集体改变,才有可能引发永久性的变异;但这一变异有可能是进一步,也有可能是退一步。
这一结论不仅是从基因稳定性理论得来的逻辑性推理,而且也是基于大量的观察得来的:某群物种每次被选择时,一开始的变化都会很快,但随后会渐渐缓慢下来,不久便停止了变化;此时,该物种会表现出与原群中少数个体所表现出的某种极端类型相同或相似的状态。
以上是从杂合子基因的污染和选择学说出发,对基因的稳定性所做的检测。关于躯体本身可能影响基因组成这一点,只是略有涉及。如果基因受到杂合子躯体性状的影响,那么,孟德尔第一定律基本假定的杂合子体内基因的精确分离,就势必不可能了。
这一结论,使我们得以直面拉马克的获得性遗传理论。在此,我们不去考虑拉马克理论的各项主张,以免离题太远,但请容我再次提及某些关系。这些关系是在拉马克学说所假定的——如果生殖细胞会受躯体影响,也即在一种性状上的改变,或许会引起某些基因内部发生相应的改变——情况下,期望得到的那些关系。现在以几个例子来阐述主要事实。
当一只黑兔与一只白兔交配时,所得杂合子幼兔为黑毛,但这只杂合子的生殖细胞内含等量的白毛基因和黑毛基因。即使所得杂合子为黑毛,也不会影响其产出含白毛基因的生殖细胞。且不管白毛基因在杂合子黑兔中停留多长时间,白毛基因仍旧为白毛基因。
现在,如果将白色基因看成某种实体,且假定拉马克学说成立,那么携带这一实体的个体,就会表现出一些躯体性状的效应。
如果将白色基因看作黑色基因的缺失,那自然就谈不上杂合子的黑色基因会对根本不存在的东西(这里指的是白毛基因)产生影响了。不过,对于信奉存缺理论的人[5]来讲,用这一观点来反对拉马克学说,是不能让他们信服的。
另一案例或许更能说明问题。让白花紫茉莉和红花紫茉莉杂交,会得到中间型的粉花紫茉莉杂合子(如图5)。如果我们将白花这一性状的出现解释为基因缺失的结果,那么红花性状的出现,必定是存在某种基因的结果。杂合子的粉红花色要比红花花色浅一点,如果性状可以影响基因,那么杂合子中红色花色的基因应该会被粉色花色的基因稀释。但在这里,或是在其他实验中,都没有记载过这样的效应。所以红色基因和白色基因在粉色花朵的杂合子中,应该是分离的,对躯体并没有任何影响。
或许,另一证据会更有力地驳倒获得性遗传理论。有一种果蝇,出现了被称之为不整齐腹缟(腹部黑缟不规则)的性状,其腹部原本整齐排列的黑缟或多或少地消失了一部分(如图152)。当培养基中的食物充足,且培养基保持较高的湿润度和酸性时,出现的第一只果蝇,其腹部黑缟损失较多。随着时间的流逝,培养基变得越来越干燥,所培养出的果蝇在外形上也越来越正常,直到后来,我们已不能将这些培养基中的果蝇和野生型的果蝇区别开来。这里,我们遇到了一个对环境影响极其敏感的遗传性状。这一性状,为我们研究躯体对生殖细胞可能产生的影响提供了机会。(https://www.daowen.com)
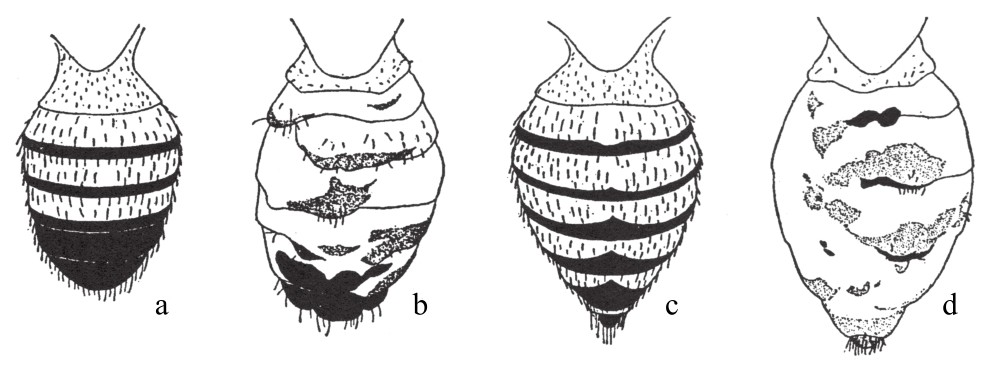
图152
图a为正常雄蝇的腹部黑缟;图b为异常雄蝇的腹部黑缟;图c为正常雌蝇的腹部黑缟;图d为异常雌蝇的腹部黑缟。
如果让早期孵出的果蝇[6]和异常黑腹果蝇交配,同时用后期孵出的果蝇[7]和正常黑腹果蝇交配,在两者交配所得的子代中,会得到相同的果蝇。最先孵化出来的果蝇的腹部是异常的,之后孵化出来的果蝇的腹部是正常的。对生殖细胞而言,祖代的腹部正常与否,是没有一点区别的。
如果有人说,第一代的影响太小,以至于看不出来变化,那么请容我补充一句,将孵化较晚的果蝇再连续繁殖十代,所得结果也不会与前面的结果有任何出入。
还有一个例子,同样能证明这一点。在果蝇中有一种突变型果蝇,被称为无眼果蝇(如图30)。这些果蝇的眼睛,比正常眼睛要小,而且变化很大。通过选择所得到的清一色的原种类型中大多数果蝇没有眼睛,但随着培育时长的增加,培养皿中将会出现越来越多的有眼果蝇,而且眼睛也越来越大。现在,如果我们将后期孵化出来的果蝇繁育起来,所得子代和无眼果蝇的子代是一样的。
这里,晚期孵化中出现的果蝇,有眼是一个更为明显的性状,可能被认为是比不整齐腹缟的例子更好的证据。在不整齐腹缟果蝇的例子里,晚期孵化出的幼虫的对称性和色素沉淀,都不是很明显的存在性状。然而,两例的结果是一样的。
近些年来,有人自以为拿到了获得性遗传的大量“证据”,但我们没有必要去逐一验证。在此,我只选取最完备的一个例子加以说明,因为该例给出了这一结论所依据的数据和定量的资料。我指的是迪肯(Dürken,1906—1984)[8]最近做的实验。这一实验似乎进行得很仔细,而且对迪肯来说,这一实验似乎还为获得性遗传提供了一个有力的证据。
迪肯用普通甘蓝蝴蝶[9]的蝶蛹做实验。自从1890年以来,人们就知道当一些蝴蝶的毛毛虫化蛹之际(当这些毛毛虫转变为静止不动的蝶蛹时),环境或照射光线的颜色,都会在一定程度上影响蛹的颜色。
例如,如果甘蓝蝴蝶的毛毛虫在白昼甚至在弱光下生活和转化,所得的蝶蛹就会稍显暗黑;但如果甘蓝蝴蝶的毛毛虫在黄光或红光下,或者在黄色或红色的帘布后生活和转化,所得的蝶蛹就会是绿色的。蝶蛹之所以呈绿色,或许是由于表层黑色素的缺失。当黑色素缺失时,内部的黄绿色会透过蝶蛹的表皮显现出来(如图153)。
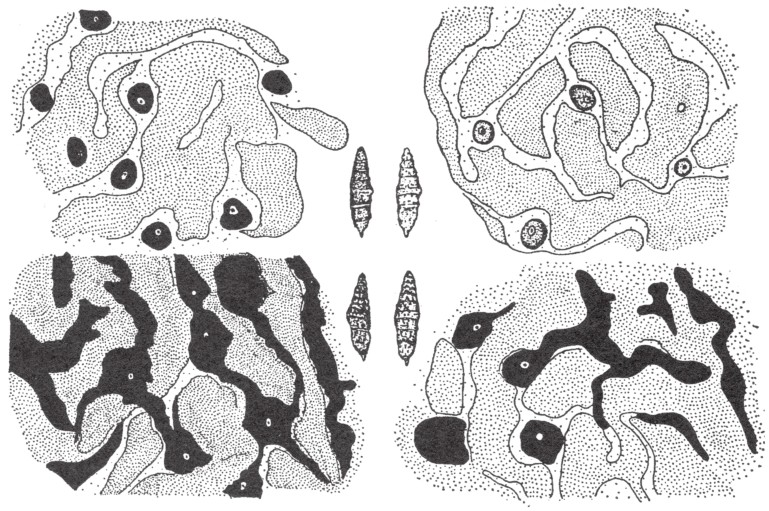
图153
图示中央为甘蓝蝴蝶四种不同颜色的蝶蛹。围绕在它们周围的,是各自的表皮层色素细胞的特殊排列。
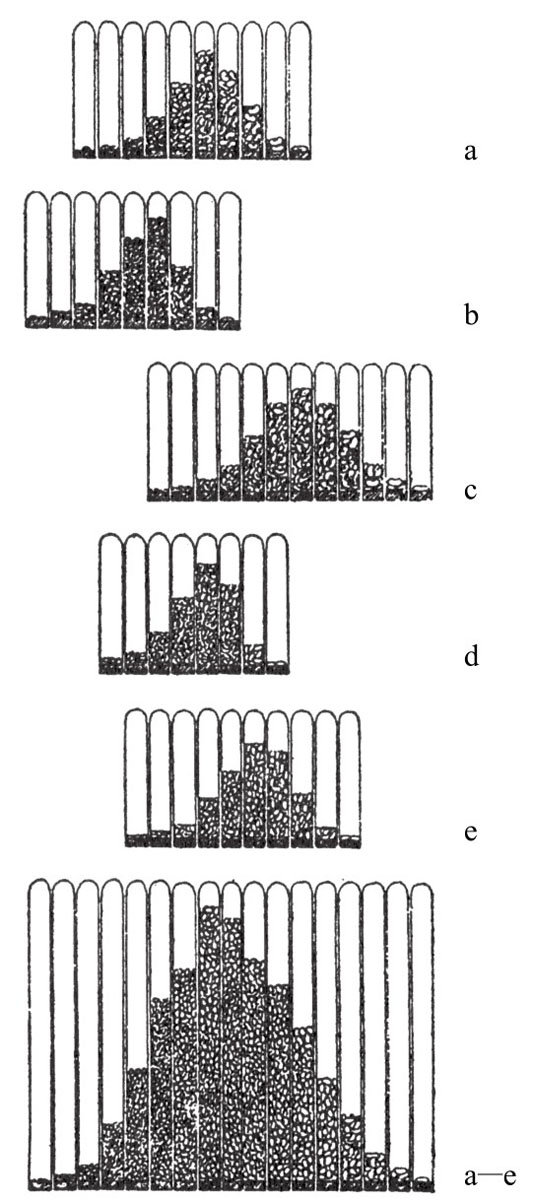
迪肯的实验是将毛毛虫置于橙光或红光下培养,得到浅色或绿色的蝶蛹。迪肯将得到的蝶蛹置于野外笼内饲养,且将这些蝶蛹所产的卵收集起来。下一步,他把由这些卵孵化而来的幼虫,一部分置于有色光照下再次培养,另一部分置于强光或是黑暗环境中再次培养。后者作为对照组,实验结果如图154所示。图中用黑色条表示暗黑色蝶蛹的数量,用浅色条表示绿色或是浅色蝶蛹的数量。事实上,蝶蛹可分为五种颜色群,其中三群被归于暗色一类,另外两群被归于浅色一类。
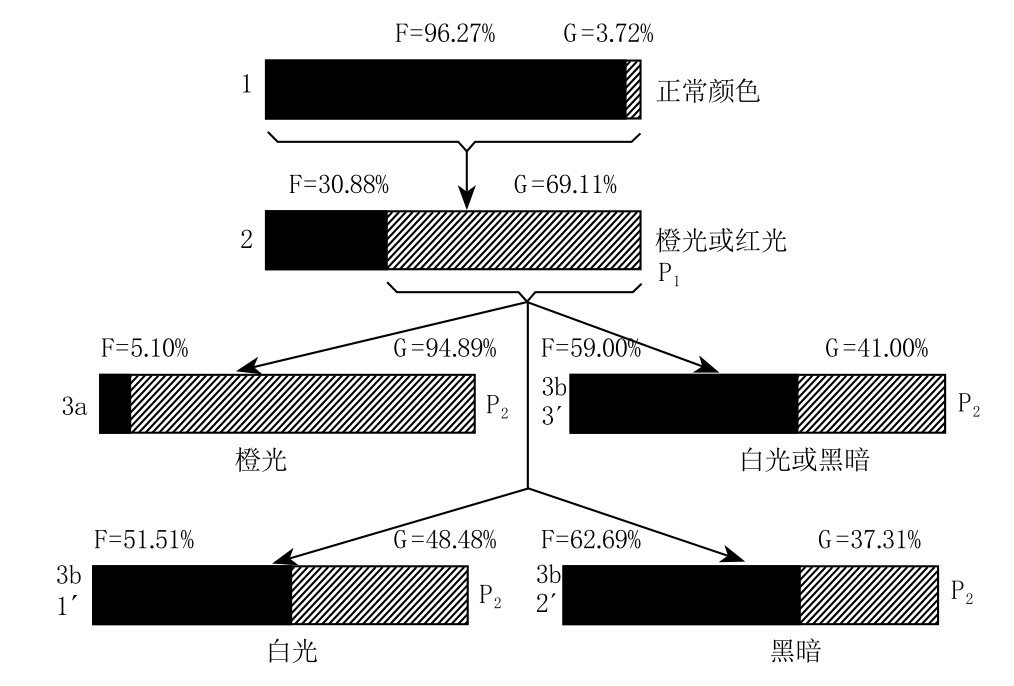
图154
甘蓝蝴蝶中,黑色蛹和浅色蛹的选择结果。
正如图154中的1所示(代表正常的颜色),几乎所有随机收集或在正常环境中收集的蛹都是暗黑色的,只有一小部分是绿色或浅色的。将这些蛹的毛毛虫置于橙光环境中饲养,当毛毛虫化为蝶蛹时,会出现占比很高的浅色类型,如图154中的2所示。如果这时只选浅色类型的蝶蛹来饲养,一部分置于橙光下饲养,一部分置于白光下饲养,还有一部分置于黑暗中饲养,那么所得结果就会如154中的3a和3b所示。3a中的浅色蛹比之前的多,因为连续两代都置于橙光中饲养,浅色的效果得到加强。然而,3b这一组的意义更为重大。如图154所示,在白光或是黑暗中饲养的蝶蛹,比野生型中的浅色蝶蛹多。迪肯认为浅色蝶蛹的增加,一部分归于前一代从橙光中继承而来的效果,另一部分归于新环境所产生的反方向效果。
目前,从遗传学的角度来看,这一解释并不是那么令人满意。这个实验表明并非所有的毛毛虫都对橙光有反应。如果对橙光有反应的毛毛虫具有不同的遗传性,那么当选用它们即实验中的浅色蛹做橙光实验,且把对照组置于白光或黑暗中培养时,我们其实已经是在处理一个反应较强的类型,即一群经过选择的类型,并且预测该种群的下一代会再次有反应,事实上也确实如此。
因此,除非一开始便选取遗传上同质的毛毛虫,或者选取其他对照组进行实验,否则,这一证据显然不能证明环境的遗传效应。
几乎所有这类研究,都犯过同样的错误。如果现代遗传学不能得出更多的成就,它也可以借展示这一证据的毫无价值来证实它自己。
现在,我们或许得转移到下一组例子。在部分实验里,有的可能是生殖细胞在经过特殊处理后受到直接损害,而受到损害的生殖物质会遗传给后代。由于生殖物质受损,后代中可能会出现畸形。这就意味着,上述特殊处理并没有通过首先使胚胎残缺来影响生殖物质,而是同时影响了胚胎及其生殖细胞。
在酒精对豚鼠的影响问题上,斯托卡德(Stockard)做过一系列实验。他把豚鼠置于充斥着高浓度酒精的密闭箱中。这些豚鼠吸入含酒精的空气,几小时后便会完全失去知觉。这一实验的时长相当久。有的豚鼠是在处理中(即失去知觉时)交配,有的豚鼠是在处理后(即苏醒之后)交配,不管是在哪种状态下交配,其所繁育出的豚鼠后代都是一样的。很多胎儿要么流产,要么被吸收,也有的一生下来就死了,或出现畸形,尤其是神经系统和眼部畸形(如图155)。只有不表现出缺陷的豚鼠,才能继续繁育后代。但在它们所繁育出的后代中,畸形的幼龄豚鼠和表面看上去正常的幼龄豚鼠相继出现。在更后面的世代中,依然会出现畸形豚鼠,但它们只会从确定的个体中产生。
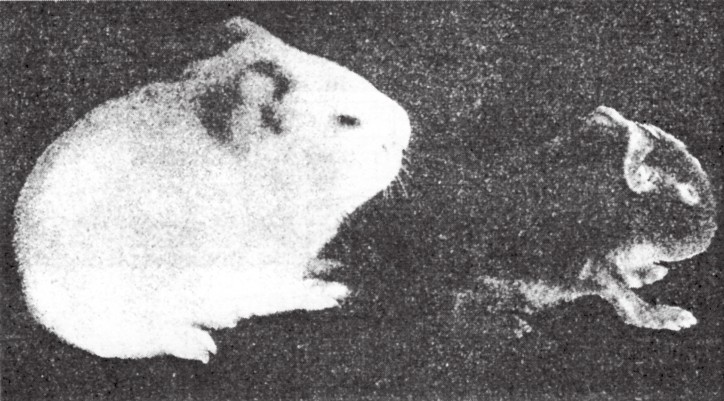
图155
两只异常的幼豚鼠,其祖先都是进行过酒精实验的。
如果我们对受酒精影响的这一豚鼠谱系加以检查,看不出实验结果与任何孟德尔式比例的证据相符合。然而,受影响的部位所表现出来的异常,也不像在单个基因发生改变时所看到的那样。另一方面,这些缺陷与我们在实验胚胎学中所了解到的用有毒的介质去处理卵细胞而导致的卵细胞发育异常,有很多共同点。斯托卡德的实验引起了人们对这类关系的关注,人们认为他的实验结果意味着酒精会对一些生殖细胞产生一定的损害——有关遗传机制某一部分的损害。之所以这些损害只对躯体中的某些部位起作用,是因为这些部位对于任何脱离正常发育轨迹的变化最为敏感。这些敏感部位多为神经系统和感觉器官。
近期,利特尔(Little)和贝格(Bagg)在妊娠鼹鼠和老鼠身上进行了一系列实验,借以研究激光的作用。经适当处理,妊娠鼠子宫内的幼鼠胚胎可能出现畸形发育。在产前检查中发现,幼鼠的脑脊髓或其他部位(尤其是四肢原基)会局部出血(如图156)。在这些胚胎中,会有部分胚胎在产前死亡,并且被吸收;还有部分会流产;但仍然有一些有活性或是得以侥幸存活下来的胚胎,可以继续繁育。这些存活下来的幼鼠后代的脑部或四肢往往有缺陷。其后代中,可能会出现无眼幼鼠,或是只有一只眼睛的幼鼠,且这只眼睛比正常的眼睛要小。贝格让这些鼹鼠进行交配,他发现其后代所表现出来的缺陷,大体上和原始胚胎中出现的缺陷相似。
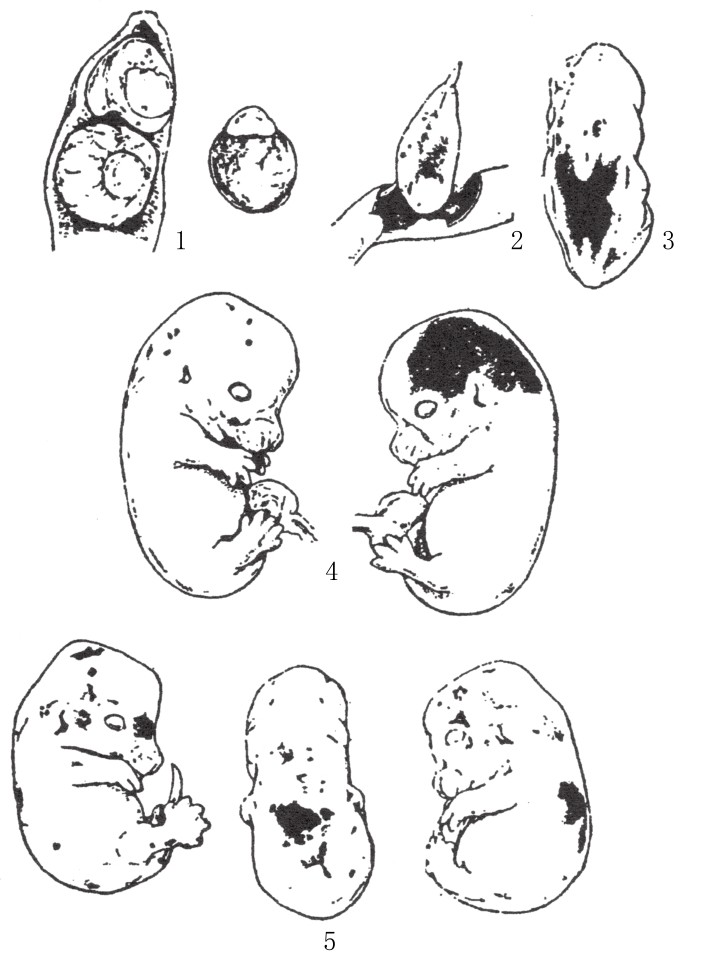
图156
当幼鼠胚胎处于妊娠鼠子宫内时,对妊娠鼠进行激光处理,其幼鼠胚胎出现局部出血。
对于这些实验,我们该作何解释呢?是不是因为激光最先对发育中的胚胎的脑髓产生了影响,从而导致缺陷,然后由于这些脑髓缺陷的存在,同一胚胎中的其他生殖细胞也随之受到了影响呢?很明显,这一解释行不通。我们能想到,当只有脑髓受到影响时,下一代预计会出现脑方面的缺陷;当眼睛是受影响的主要器官时,下一代中预计会出现眼睛方面的缺陷。然而截至目前的所有报道,都没有出现类似的情况。有着正常眼和畸形脑的鼹鼠,可能生育出有缺陷眼的鼹鼠。换句话说,特定效应是不存在的,有的只是一般效应。
还有另一种解释:激光会影响到子宫中幼鼠胚胎的生殖细胞。那么,从这些生殖细胞发育而来的新世代个体,都会有缺陷,因为在正常发育过程中最容易受到干扰的器官,也就是最容易被发育过程中的任何改变所影响的器官。总之,这些器官是最脆弱的,或者说它们是发育中平衡阶段最微妙的,因此这些器官最先表现出异常(即正常发育的脱轨现象)。我认为,这是目前对于这些实验或是其他类似实验最合理的解释。
[1]豚鼠:又名天竺鼠,无尾啮齿动物,身体紧凑,四肢短小,头大颈短,具有小的花瓣状耳朵,其耳朵位于头顶的两侧,具有小三角形嘴。
[2]1909年,丹麦植物学家约翰森将遗传因子命名为基因。
[3]正向突变:正常存在于某物种中的一个野生型基因A,可以突变为隐性基因a,这个过程一般称为正向突变,或正突变;反之,隐性基因a也可以变成野生型基因A,这称为回复突变,或反突变。
[4]拉马克学说:由法国博物学家拉马克提出的关于生物进化的系统看法。主要内容是:
(1)认为地球有悠长的历史,决非像特创论者所说的那样只有几千年历史,而且地球表面不是固定不变的,而是经历了一个不断渐变的过程。
(2)认为生命物质与非生命物质有本质区别;生命存在于生物体与环境条件的相互作用之中;低级生物类型可以不断地由非生命物质自然发生出来;植物和动物虽有重大区别,但有共同的基本特征;生命即运动,运动表现在各方面,既表现在生物体内液体的流动上,也表现在生物体吸收养料和排出废物上;生命是连续的、变化的、发展的。
(3)认为物种之间是连续的,没有确定的界限,物种只有相对的稳定性;物种在外界条件影响下可能发生变异,栽培植物和饲养动物的出现就是物种变异的例证;古代物种是现代物种的直接祖先,物种一般不会消灭;动物界普遍有种间斗争,种内斗争则不常有。
(4)认为生物进化的动力,一是生物天生具有向上发展的倾向,这是生物向上发展的原因。二是环境条件的变化,环境条件的改变会引起生物发生与之相适应的变异,环境条件变化的大小,决定着生物发生变异的程度;环境条件的多样性是生物多样性的原因。
(5)认为在植物和低等动物中,环境的改变会引起功能的改变,功能的改变会引起结构的改变;而在具有神经系统的动物中,环境的改变先引起生活需要的改变,生活需要的改变又引起习性的改变,新习性的发生和加强,引起身体结构的变化;凡经常使用的器官会发达进化,而经常不用的器官就会萎缩退化(即用进废退),这些后天获得的性状能够遗传给后代(即获得性遗传),这样经过一代代积累,就会形成生物的新类型。
(6)认为无论植物或动物,都按一定的自然顺序进化,由简单到复杂,由低级到高级;进化是树状的,即不只向上发展,也向各个方面发展;人类大概是由高级猿类发展而来的。
(7)最原始的生物源于自然发生。拉马克支持生物进化多元论,生物有多个祖先。
拉马克学说曾在科学界发生过重大影响,为以后生物进化论的发展奠定了基础。但是,由于当时生产水平和科学水平的限制,拉马克对进化原因的解释过于简单化。“生物天生具有向上发展的倾向”缺乏物质基础;“环境的改变会引起生物发生与之相适应的变异”也缺乏事实根据;“器官用进废退”在当代是可能的,但这种后天获得的性状,如没有影响到遗传物质,是根本不能遗传给后代的。
[5]这里指的是那些将白色基因看作是黑色基因缺失的一派人。
[6]即处于高酸性和湿润环境下,腹缟高度不整齐的果蝇。
[7]即处于低酸性和干燥环境下,腹缟高度整齐的果蝇。
[8]迪肯:英国海洋学家,早年曾在伦敦皇家学院攻读化学。
[9]甘蓝蝴蝶:又称为菜粉蝶,别名菜白蝶,幼虫又称菜青虫,是我国分布最普遍,危害最严重,经常成灾的害虫。嗜食十字花科植物,特别偏食厚叶片的甘蓝、花椰菜、白菜、萝卜等。在缺少十字花科植物时,也可取食其他寄主植物,如菊科、白花菜科、金莲花科、百合科、紫草科、木犀科等植物。