低等植物的性别及其定义
近年来,在伞菌[17]或担子菌[18]的研究中,雌雄性别的名称问题显得尤为突出。根据汉纳(Hanna)最近的陈述,在伞菌中,“真菌学家对性别问题的关注已经长达一百多年了”。邦索德(M. Bensaude)女士(1918),克内谱(Kniep,1919—1923),芒斯(Mounce)女士(1921—1922),布莱(Buller,1924)以及汉纳(1925)的发现,都揭示了一个有趣的现象。为了行文的简洁明了,这里只详论汉纳最近的文章。汉纳运用新的精细手法,从伞菌的菌褶中分离出单个孢子,再将所得孢子置于粪胶培养基内。这样一来,每个孢子都会发育成一株菌丝体[19]。再让这些单孢子型的菌丝体一株一株地彼此接触,便能知道每一株的性别。在这些组合中,某些会彼此连合在一起,并形成长着“锁状连合”的一株二级菌丝体,由此表明这两个接触的菌丝体有着不同的性别。之后,这样的二级菌丝体会发育出子实体或伞菌。如果其他组合搭配在一起,却不会形成长着“锁状连合”的二级菌丝体,也不会发育出子实体和伞菌,因此,汉纳把这种接触在一起的两个菌丝体视为同一性别。
现在,对源于相同品系(生长在同一地区的植株)的单孢子型菌丝体加以鉴定,所得结果如表11所示。此处,用“+”表示两株单孢子型菌丝体接合后会呈现出锁状连合(两者为不同性别),用“-”表示不会呈现出锁状连合(两者为相同性别)。图表中,我们将菌丝体分为四类(属于相同组的菌丝体被安排在一起)。这一结果被解释为,这里所研究的墨汁鬼伞菌[20]中一个子实体中的孢子,属于四个不同的性别群。
表11 相同品系墨汁鬼伞菌孢子的四种性别群
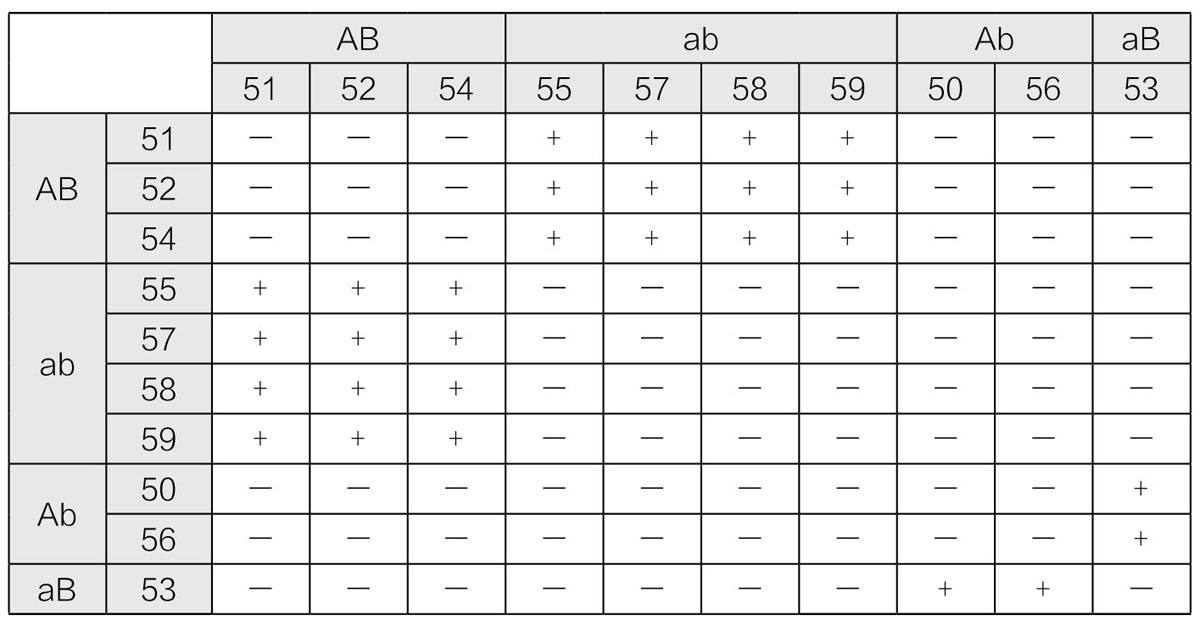
就像克内谱首次展示的那样,上述四群能用两组孟德尔式因子的假设来加以解释,即Aa和Bb。如果将这四个因子分离,当每个担子[21](也称为孢子台)形成孢子时,伞菌中将会出现四种孢子,即AB、Ab、aB和ab。每一种孢子会产生具有相同基因组合的菌丝体。如表11所示,只有那些由不同因子组合而形成的菌丝体,才会形成锁状连合。这意味着,存在四种性别,且只有那些性别因子不同的菌丝体才能连合起来。
还有一个细胞学背景,与这些遗传学假设高度吻合。在单孢子型菌丝体的细胞质中,有很多细胞核。在两条菌丝体连合之后,衍生出来的菌丝体(次生菌丝体[22])内的细胞核是成对存在的。我们有理由假设,在成对的两个细胞核中,一个来自一条菌丝体,另一个来自另一条菌丝体(即分别来自两条菌丝体)。当四个孢子即将发育成形时,它们会发生减数分裂,以至每个孢子都含有分裂后的细胞核。所得的每个孢子,都会发育成新的减数菌丝体。相同的场景也发生在高等植物和动物的减数分裂过程中,因此,这些霉菌同二倍染色体减少至单倍型配子时所发生的遗传学结果相一致。当然,墨汁鬼伞菌和其近亲种系中这种二倍—单倍的关系尚未得到证明,但这似乎不太可能是对已知事实的正确解释。如果真是如此,那么遗传因子在伞菌中的分离,原则上就和其他植物或动物是相同的。
上述关系,适用于同一地区的各个品系之间。如果检测不同地区的品系,也会得出非常相似的结果。任意一种品系的单孢子型菌丝体,都可以和其他品系的所有单孢子型菌丝体连合起来(得到有锁状连合的菌丝体,等等)。在表12中的数据,来自同一个地区(加拿大埃德蒙顿)子实体的11对单孢子型菌丝体,与来自另一地区(加拿大温尼伯)的一个子实体的11株单孢子型菌丝体的连合实验。无论何时,让来自不同区域的各品系互相交配,都会有相同的结果。汉纳所得出的组合中,墨汁鬼伞菌有20种性别。如果我们再将其他地区的伞菌囊括进去,那性别的数目必定会增加。
表12 不同品系墨汁鬼伞菌孢子的性别群
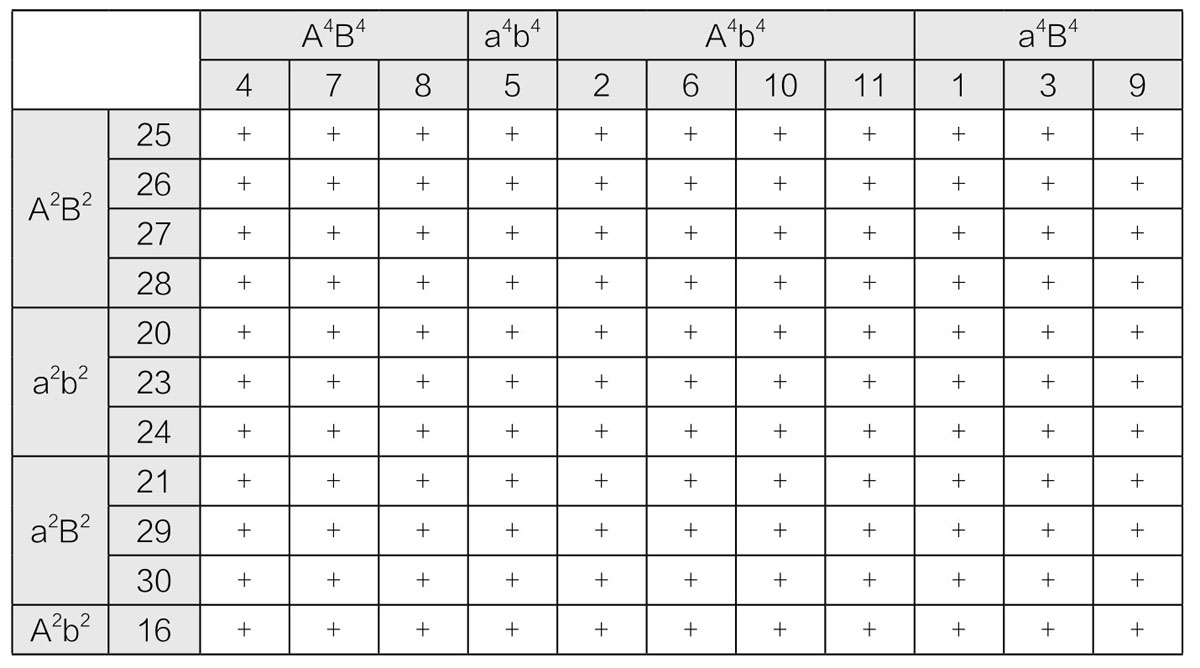
汉纳不仅进行了种系内的杂交,还用杂交过的品种来做一系列实验,进一步验证因子假说。如果将源自不同品系的因子当作成对的等位因子,用Aa和Bb来表示一个品系上的因子,用A2a2和B2b2表示另一个品系上的因子,那么,这两个变种的菌丝体结合后,将会得到16种杂合子;且每株杂合子菌丝体的行为方式,也将和纯种菌丝体的行为方式相似,即只有携带不同因子的两株菌丝体,才能形成锁状连合。
如果我们从习惯的意义上来理解有关因子,那么,在此便有着规模广泛的两性现象了。如果以这样的基础去定义性别是有益的,那么采用这一表述便不会引起反对。就我个人看来,采用伊斯特关于烟草研究的解释模式,把有关因子称为“自交不育因子”(见下文),或许那样会更简单。不论大家倾向于用哪些文字来描述,原则上解释都一样。
在《相对的性别之研究》这本书中,哈特曼(Hartmann)对他从海藻(长囊水云)上观察所得的研究结果作出如下描述。这些从植株中游离出来的自由移动的游动孢子[23]是极其相似的,但根据其后期的行为,我们将其分为两类:“雌性”和“雄性”。前者(雌性)很快就会停顿下来,然而后者(雄性)会成群地继续游动一段时间,并且围绕着雌性个体运动(如图138)。
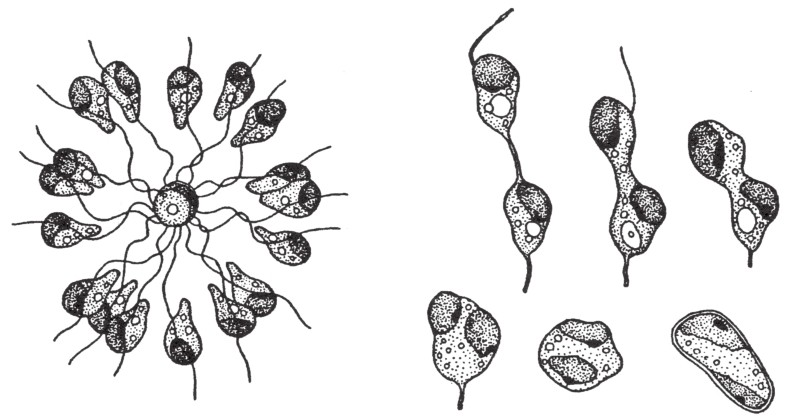
图138
左侧展示了一群雄性配子围绕着一个雌性配子;右侧展示了雄性配子和雌性配子的结合过程。
一个雄性游动孢子会与一个静止中的雌性游动孢子融合。哈特曼将祖代植株一个个孤立出来,并在这些植株释放出游动孢子时,集体测试这些不同植株孢子间的相互关系。
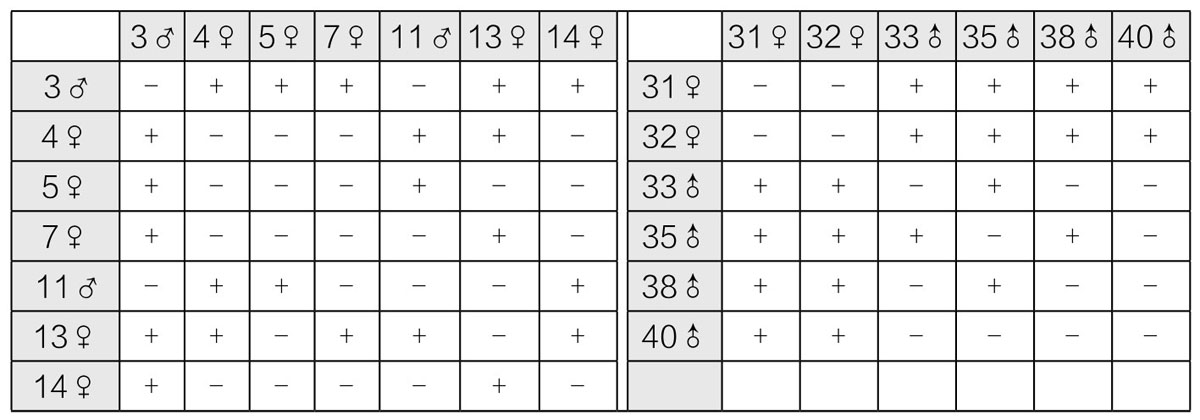
典型实验结果如表13(左侧),若游动孢子接合在一起,用“+”表示,若游动孢子不能接合在一起,用“-”表示。检测时,每一种孢子的个体都用其他种类的孢子个体进行逐一检测。大多数情况下,某植株释放出的游动孢子对于其他种类的孢子来说,要么一直表现为雄性,要么一直表现为雌性;但游动的孢子在一些个别案例的接合中表现为雌性作用,在另一些案例的接合中表现为雄性作用。例如4号(表13,左侧)和13号接合所得出的结果,就和这两者分别与其他孢子接合所表现出的雌雄作用不同。我们还在35号和38号之间的反应中发现了例外(表13,右侧),两者与其他孢子接合之后,对于与其接合的孢子表现出雄性作用,但两者相遇时,却又分别表现出雌性和雄性作用。哈特曼根据不同接合方式形成的“群”数,指出某些个体可能会呈现为强雌性,而其他个体呈现为弱雌性。他还得出结论:弱雌性相对于强雌性起雄性作用,相对于强雄性起雌性作用。这些强弱相对的关系,究竟受年龄因素(例如静止下来)或环境因素有多大程度的影响,目前还不十分清楚。虽然哈特曼对游动孢子进行检测时发现孢子天天保持着这些关系,似乎排除了这一解释,但不幸的是,该材料(游动孢子)不适合用来作为所含因子的遗传分析。是否某个体中释放出来的配子很快静止下来,就足以鉴定性别?如果是,那么弱雌性又是如何表现为雄性作用的?诸如此类问题,也是不明确的。不过同一植株的配子不能交配,这一现象似乎属于自交不孕现象和杂交可孕现象的范畴。当前,如果将其作为判断性别的标准,很大程度上得依赖于个人的选择和定义。就我个人而言,如果将“性别”一词用于一个配子与另一个配子结合与不结合的现象,而不是用于普通的所谓性别现象,那么,不但不能阐明有关问题,反而更容易混淆性别一词所关涉的问题。
表13 游动孢子接合实验的结果
现在我们不妨提出这样一个问题,将有关因子称为自交不育因子,而不是性因子,是否会更简单且不那么容易引起困惑呢?伊斯特最近关于烟草自交不育研究的重大成果,为研究多次的显花植株中杂交可育以及自交不育的问题,第一次奠定了证据确凿的遗传学基础。显花植株中的这些现象,与墨汁鬼伞菌的菌丝体和长囊水云的游动孢子的接合十分相似,虽然两者在过程方法上不完全相同,但其遗传学和生理学背景可能是基本上相同的。
伊斯特和曼格尔斯多夫(Mangelsdorf)花了几年的时间,研究两种烟草杂交中的自交不育问题。在一篇简短的报告里,他们对这一研究做了总结。在此,我们只给出最一般性的结论。他们运用特殊的操作方式,将自交不育的个体培育成了几种自交且纯合的品系,可以稳定持续12个世代之久,从而得到了用于测试这一问题的合适材料。在此,只给出品系中一种类型所产生的结果。有三类个体a、b和c,任何一类的每个个体与同类的其他个体交配时,都表现出自交不育,而与其他两类的个体交配时,却是可育的。但通过正交和反交得来的后代有所不同。由此,让雌性a个体与雄性c个体交配,只会得到b个体和c个体;而让雌性c个体与雄性a个体交配,只会得到a个体和b个体。所得后代中的两种个体的数量各占一半,但后代中都没有出现母方的那类个体。对于这一点的解释如下:如果在此种系中存在等位基因S1S2S3,且a类的基因为S1S3,b类的基因为S1S2,c类的基因为S2S3,加上如果一植株的雌蕊柱头只能刺激其他有着不同等位基因的花粉,而对有着和自身相同的基因的花粉不起作用,那么这个结果就能解释得通了。例如,c植株(S2S3)只会对携带S2和S3以外的基因的花粉产生足够的刺激,既只有携带S1因子的花粉才能进入c植株并对其卵细胞进行受精。其得到的后代会是同等数量的S1S2(b类)和S1S3(a类)。将其反交时,即用雌性a类(S1S3)个体与雄性c类(S2S3)进行交配时,S2因子会独自进入a类的卵细胞内,得到S1S2(b类)和S2S3(c类)。这一结果对于其他类来说也是适用的,这就解释了为什么在其后代中不会得到母方那类个体,为什么正反交所得后代是不同的,以及为什么不论父方是哪一类,后代中两类个体(没有母型)的数量是相同的。
有很多种方法,可以验证这个假设的真实性。而我们所做的验证,也证实了这一假设。这一令人信服的分析,是经过精心设计而得出的遗传实验的结果,对困扰了学者们超过75年之久的问题来说,确实是头等贡献。这一解决办法不仅对本案例进行了精辟的基因分析,而且深入透析了单倍型花粉管和二倍型雌蕊组织的生理反应。通过直接观察可知,雌株组织中的花粉管的生长率[24]是符合差等生长率确实存在的这一观点的。目前,这种关系的本质是什么我们还不知道,但可以合理假定它是化学性的。极有可能,我们可以利用相似或相同的化学反应及其遗传学基础来解释:为何在低等植株中不同的菌丝体在连合时会有自交不育的现象。如果这一说法能够成立,那么,遗传学问题的主要研究对象,首先就与自交不育因子相关,而这些因子极有可能是孟德尔式因子。把这些因子定义为性别因子,至少是通常适用于雌雄异体的生物躯体上的性别因子,似乎是很值得怀疑的。的确,在这些差异里,也存在产生以互相连合为主要机能的精细胞和卵细胞的那些差异,但是就一般理解而言,这些机能和那些雄性和雌性个体在体质上所体现出来的机能相比,还是不那么显而易见。
[1]根据麦克朗(McClung)所说,雄性蚱蜢的X染色体不一定都一成不变地附着于特定的常染色体上。尽管在某一个体中的X染色体的附着是恒定的,但在其他个体中,X染色体的附着是自由的。如果此类型中的这类性状是性连锁的话,那么它们的遗传可能是相当复杂的,毕竟X染色体和常染色体之间的附着关系不恒定。
[2]即父母的性状和原来实验时父母的性状完全相反。
[3]有可能在451次中出现1次X染色体与Y染色体的交互。
[4]即染色体之间发生交换而得到的杂合子类型。(https://www.daowen.com)
[5]另一个实验中,交换型占雄鱼的1/17。
[6]即温格用MmEe来表示染色体上的基因公式。
[7]瘤蚜:一种葡萄树害虫,可毁坏葡萄树的根部。
[8]蚜虫:又称腻虫、蜜虫,是一类植食性昆虫,包括蚜总科(又称蚜虫总科)下的所有成员。目前已经发现的蚜虫总共有10个科约4 400种,其中多数属于蚜科。
[9]所以这个瘤蚜雌虫被称为成雄卵细胞之母。
[10]在某种轮虫中,两种大小的卵细胞都是由雌性产生的。两种卵细胞都会分裂出两个极体,从而形成单倍型原核。两种卵细胞在受精之后,稍大的卵细胞会发育成为雌性,较小的卵细胞会发育成为雄性。目前,为什么卵巢会产生两种大小的卵细胞,我们还完全不知道。
[11]线虫:线虫动物门是动物界数量最丰者之一,除少数自由生活于土壤、淡水和海水环境中,绝大多数营寄生生活,不过,只有极少部分寄生于人体并使人致病。常见的线虫有蛔虫、鞭虫、蛲虫、钩虫、旋毛虫和类粪圆线虫。
[12]休眠卵:在动物繁殖的过程中,为了应对恶劣的环境,一些动物的胚胎外面会被一层包被膜包裹,此时,动物的胚胎发育处于休眠期,等到条件合适的时候再发育,这样的动物胚胎就叫作休眠卵。
[13]当没有受精的成雄卵子进行分裂时,每条染色体都会断裂成两部分(形成种系的细胞核可能例外)。这一过程并未出现染色体的分裂,而是出现染色体断裂或分开成两个片段。如果这一解释能说得通,那么基因不会有数目上的增加,这一过程的出现(线虫中也出现过)也不会对性别决定机制有任何影响。
[14]根据怀丁所说,“黑眼偏父遗传的雄蜂,在形态畸变的占比上,比正常产出的雄蜂和雌蜂的更高。大多数偏父遗传的雄蜂,已被证明是没有生殖力的;个别黑眼雄蜂,有部分生殖力;此外,还有少数嵌合体产生了橘眼雌性后代,这些后代有完全的生殖力。在偏父遗传的雄蜂的下一代中,橘眼雌蜂在形态上和可孕性上,都是正常的;而黑眼雌蜂数量较少,出现畸形的可能性极大,并且其几乎没有生殖能力。”寄生蜂中的特种雄蜂,可以说明蜜蜂中出现的一些不规则情况。
[15]属于苔藓植物门,苔纲,囊果苔目。植物体小形,倒心脏形,不分枝或两歧分枝,单层细胞,中肋宽阔,由多层细胞组成。
[16]膜翅目昆虫种类众多,其生活方式和生理结构差异极大。一般这些昆虫拥有两个透明的、膜一般薄的翅膀,翅膀上的脉将每个翅膀分为面积比较大的格,翅膀的运动方向一般相同。有些膜翅目的昆虫的翅膀已完全退化了(比如蚂蚁中的工蚁)。飞行时膜翅目的两个翅膀一般同步运动。大多数膜翅目昆虫有两个大的复眼和三个小的单眼。一般膜翅目昆虫的口器可以咀嚼,但也有一些昆虫用嘴来舔吸,比如蜜蜂。膜翅目昆虫是全变态类昆虫中唯一有产卵管的昆虫,许多膜翅目昆虫的产卵管变异为一根毒针。通常,雄性膜翅目昆虫通过孤雌生殖形成,雌性膜翅目昆虫从受精卵孵化而成。
[17]伞菌:一般指具有菌盖和菌柄的肉质腐生菌类。多数伞菌可供食用,如香菇、蘑菇、草菇等;少数种类有毒,如毒鹅膏菌、奥来丝膜菌。
[18]担子菌:由多细胞的菌丝体组成的有机体,菌丝均具横隔膜。
[19]菌丝体:菌丝的集合体,纵横交错,形态各异,具有多样性。菌丝细胞的分裂多在每条菌丝的顶端进行,前端分枝。菌丝在基质中或培养基上蔓延伸展,反复分枝成网状菌丝群,通称菌丝体。
[20]墨汁鬼伞菌:又名鬼盖、鬼伞、鬼屋、鬼菌或朝生地盖,分类在鬼伞属下,是继毛头鬼伞菌(鸡腿菇)后第二著名的鬼伞菌。
[21]担子:指的是担子菌纲所特有的分生孢子梗。它们或是无隔的(如无隔担子菌亚纲),或是分隔的或是分枝的,有时是由一个孢子或类似孢子的结构(如有隔担子菌亚纲)发育而来的。担子菌的有性孢子的数目固定(如4个),通常由双核菌丝顶端细胞膨大呈棒状的担子,经过核配和减数分裂生成4个单倍体细胞核,并在担子上生出4个小梗,4个核分别进入小梗内,最后在小梗顶端形成4个外生的单倍体孢子,称为担孢子。
[22]其功能至少有吸收营养、代谢物质的运输、代谢产物的储藏及繁殖等四种。按照发育顺序,菌丝体可分为初生菌丝体、次生菌丝体和三生菌丝体。
[23]游动孢子:从孢子结构来看,孢子可分为游动孢子和不动孢子两种类型。游动孢子是指具有鞭毛可以游动的孢子,多见于某些藻类和真菌。它既能进行无性生殖,也可以在某些条件下进行有性生殖。不动孢子是指不具鞭毛的、不能游动的、具细胞壁的孢子。
[24]花粉管形成后,花粉粒的内容物质包括精子全部进入花粉管并集中于花粉管前端,并随着花粉管的生长逐渐向前移动。花粉管生长穿过柱头,伸入花柱;通过花柱,到达子房;经过珠孔,进入胚囊。