第九章 三倍体
第九章 三倍体
由于基因间的平衡得以保持,我们预计三倍体胚胎的发育应当是正常的。唯一的不和谐因素,可能是三组染色体与遗传所得的细胞质分量两者间的关系。生物自身的调控作用,影响有多深远,我们还不怎么明确,但可以总结如下:至少在植物中,三倍体的细胞要比正常型(二倍体)的细胞大。
在近期的著作中,有大量关于三倍体[1]的记录。这些三倍体,有一部分是从已知的二倍体中产生的,有一部分是从培育植株中发现的,还有一部分是在野生状态下发现的。
盖茨和卢茨(A. Lutz)描述过有着21条染色体的三倍体待霄草(半巨型)随后,德弗里斯、范·欧沃瑞姆(van Overeem)和其他学者也对三倍体待霄草有过描述。三倍体待霄草的产生是由于二倍体与单倍体生殖细胞的融合。
盖茨、海尔茨(Geerts)和范·欧沃瑞姆分别对成熟期三倍型染色体的分布状况做了研究。他们发现,在一些例子中减数分裂时期的染色体分布很规律,然而在另外一些例子中,有些染色体却被丢弃,后期便退化了。卢茨女士发现,实际上由三倍体产生的后代有很大变异。盖茨对有着21条染色体的植物做了记录,由第一次成熟分裂所得的两个子细胞含有“几乎恒定”的10条染色体和11条染色体,而含有9条染色体和12条染色体的情况十分罕见。海尔茨发现了更多的染色体不规则分布的现象。他指出,有7对染色体通常会分别去向每一极,剩余的不成对的7条染色体会不规则地分布到两极。他的这一发现,符合7条染色体会和另外7条染色体接合,其余7条染色体找不到接合对象的情况。范·欧沃瑞姆表示,如果用三倍体待霄草做母株,结果证明,不管孤立无偶的染色体的分布如何不规则,大多数胚珠都是有用的。换句话说,所有或大多数不同的卵细胞群会存活下来,并有可能受精,结果导致许多有着不同染色体组合的各种各样的植株出现了。另一方面,如若使用三倍体待霄草的花粉,结果显示只有这些含7条染色体或是14条染色体的花粉会继续完成受精。而含中间数目染色体的花粉粒,大多不能继续完成受精。
德摩尔在栽培风信子[2]时,发现了三倍体。根据他的陈述,出于商业需求,这些三倍体风信子正在取代以前的老品种。一些风信子衍生品种含有三倍左右的染色体,它们成为了现代培育品种的重要部分。由于风信子是用球茎繁殖的,所以任何特殊的品种都能继续繁育。德摩尔对正常型风信子和三倍体风信子的成熟生殖细胞做了研究(如图80)。正常型二倍体风信子的成熟细胞有8条长染色体,4条中等染色体和4条短染色体。其得出的单倍型生殖细胞含有4条长染色体,2条中等染色体和2条短染色体。德摩尔和贝林都指出,“正常型”或许早已经是四倍体了,因为在减数分裂后所得的子细胞中,每一种长度的染色体都有2条。如果真是如此,所谓的三倍体很有可能就是双重三倍体,因为它有12条长染色体,6条中等染色体和6条短染色体。
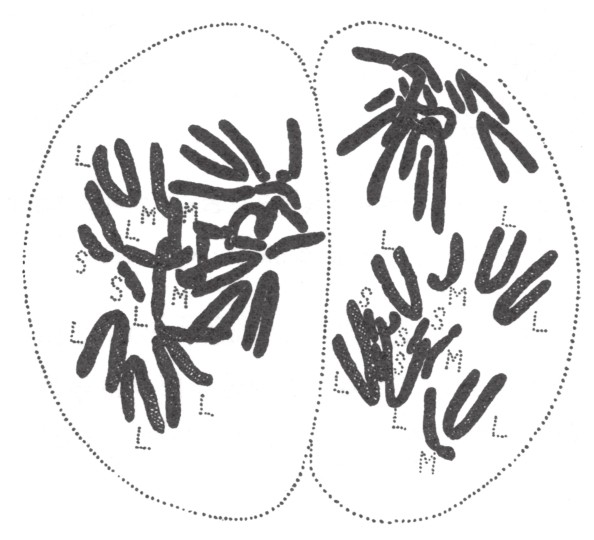
图80
风信子花粉母细胞的三倍型染色体群。
贝林也对变种三倍体美人蕉(Canna)的成熟分裂做过研究,他发现各种大小的染色体都是以三条为一组存在的。当三条染色体分离时,一般情况下,都是两条一起进入一极,另外一条进入另外一极,但因为不同类型的染色体的分布方式是随机的,以至于分裂后的姐妹细胞中只有极少数会成为一个二倍型细胞和一个单倍型细胞。
布莱克斯利、贝林和费恩曼(Farnham)报告过一种三倍体曼陀罗。它是由正常型二倍体和四倍体植株受精而得来的。正常型二倍体植株中有24条染色体(n=12)(如图81a)。三倍体植株有36条染色体(n=18)(如图81b)。单倍染色体群组成有:1条超大号(L),4条大号(l),3条中等偏大号(M),2条中等偏小号(m),1条小号(S)和1条超小号(s)。因此,二倍型染色体群为2(L+4l+3M+2m+S+s),再推之,三倍型染色体群组便是三倍这种搭配。
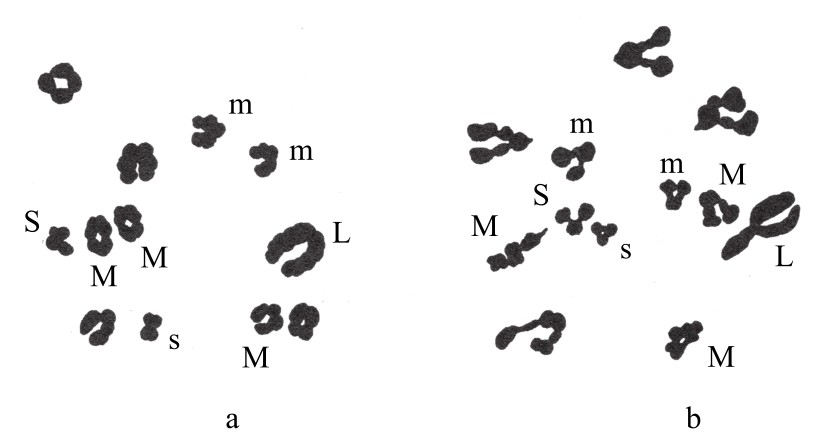
图81
图a为二倍体曼陀罗减数分裂的染色体群;图b为三倍体曼陀罗减数分裂的染色体群。
贝林和布莱克斯利对三倍体物种的成熟分裂做过研究。减数分裂后的染色体群有12组染色体,每组3条(如图81b)。这些三价染色体群间的体积比和二倍体的二价染色体群间的体积比相同,即它们都仅由相同的染色体组合而成,并以图示中不同的方式接合。例如有两条染色体两端相连,另一条染色体只有一端与其他两条相连,等等。
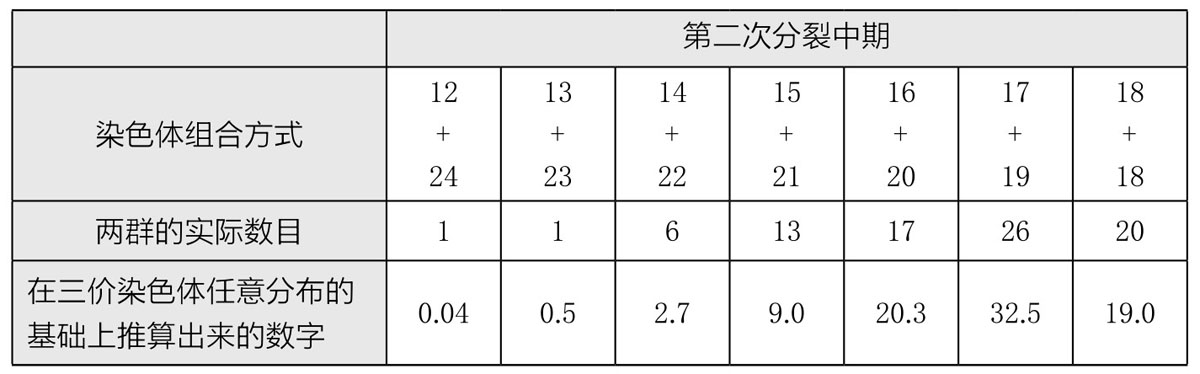
在第一次分裂时,每组三价染色体中的两条去向纺锤体的一极,另外一条去向另外一极(如图75,第3列),不同类的三价染色体再次随机组合,进而得出若干不同的染色体组合。表2所记录的,是三倍体曼陀罗84个花粉母细胞所含的染色体数目。实验结果与随机组合预计得出的数字高度吻合。
表2 三倍体曼陀罗84个花粉母细胞的染色体组合
三倍体偶尔会出现第一次成熟分裂进行不了的情形。短时间的低温处理,有利于阻碍第一次减数分裂的发生。如果在不发生第一次减数分裂的情况下,发生了第二次分裂,染色体会进行均等分裂[3],得到各含有36条染色体的两个巨型细胞。
一般情况下,三倍体所得的花粉很少能用于受精,但其卵细胞大多可用于受精。例如,用正常型植株(2n)给某三倍体植株授粉,所得的正常后代的数量会远远超过三倍体卵细胞和花粉自由授粉时预计所得的数量。
布里奇斯发现了三倍体果蝇(如图82)。三倍体含三条X染色体,它们同各类三条常染色体相平衡,所以这些果蝇都是雌性。这种平衡,是产生正常雌性的关键。既然所有染色体的遗传因子是已知的,那么,我们就有可能利用后代性状分布的情况,来研究染色体成熟期内的活动。对交换加以研究,来确定染色体是否是以三条为单位互相配对的,也是有可能的。
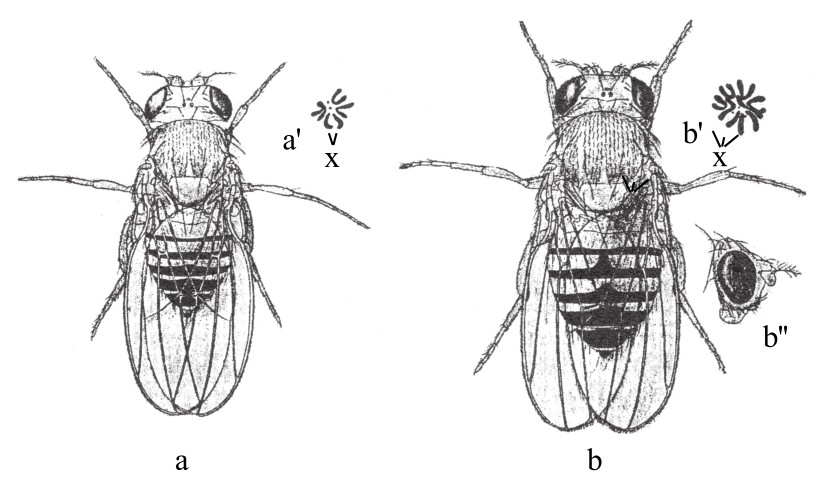
图82
图a为正常型二倍体果蝇;图b为三倍体果蝇。
真正的三倍体果蝇中,既有三组普通染色体也有三条X染色体。反之,若只有两条X染色体,则该个体显示为性中型(中间型);若只出现一条X染色体,则该个体显示为超雄性。它们的关系如下所示:
3a+3X=三倍体雌蝇
3a+2X=性中型果蝇
3a+1X=超雄性果蝇
在雌雄同体的动物中,研究者发现了另一种在胚胎时期是三倍体的物种。据相关研究,雄性蛔虫的二价型变体,其成熟的卵细胞中含有两条染色体,将这种卵细胞与只含一条染色体的精子结合,最终这些卵细胞都会发育成含三条染色体的胚胎。但这些胚胎在生殖细胞未发育成熟之前就和母体脱离了,所以,并未观察到染色体在接合时期发生融合这一重要特点,也没有发现过成熟的三倍体蛔虫。
还有一种三倍体,是通过将两个二倍体杂交,之后将其杂合子(因为接合和减数分裂没有发生,所以得到的是二倍体的生殖细胞)与亲型原种回交所得。费德莱(Federley)对三种飞蛾做过这种实验,这三种飞蛾的染色体数目如表3所示:
表3 三种蛾类的染色体数目
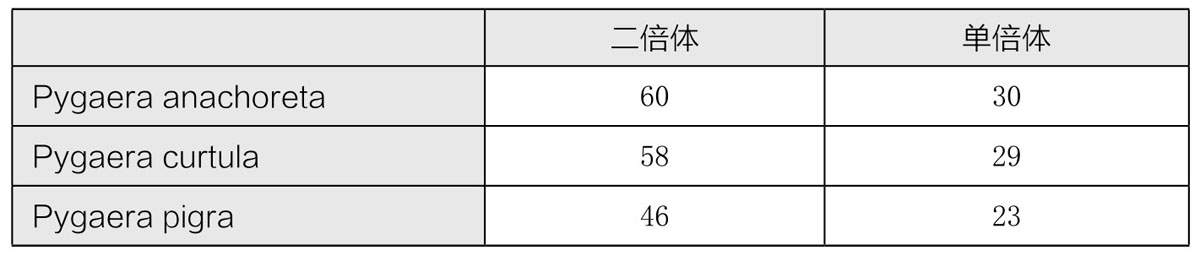
前两种蛾的杂合子有59条染色体(30+29)。当这种杂合子的生殖细胞进入成熟期时,其染色体之间不会相互接合。在第一次成熟分裂时期,59条染色体每条都会分裂到两个子细胞中去,每个子细胞都会含有相同的染色体数目(59)。在第二次减数分裂时,就会出现不规则的性状,每条染色体再次分裂成两条,但这两条染色体一般不会分离。即便如此,子一代雄性中仍有部分是可育的,正如结果所展示的那样,有一部分雄性生殖细胞内含有完整的染色体数目(59)。这样,子一代中的雌性就是不可育的。
如果子一代中的雄蛾与亲代的雌蛾回交,例如,就拿anachoreta蛾来说,其卵细胞中含有30条染色体,所得的子二代杂合子中有89条染色体(59+30),因此,这是一个三倍体杂合子。这些子二代(三倍体)杂合子与子一代(二倍体)杂合子相似。前者拥有两套anachoreta蛾的染色体和一套curtula蛾的染色体。从某种意义来讲,它们是一种永久性的杂合子,尽管在各个世代中,它们的染色体只有半数能彼此接合。例如,当含有89条染色体的杂合子的生殖细胞成熟时,两组anachoreta蛾的染色体(30+30)会彼此接合,而剩下的29条curtula蛾的染色体则保持孤立。第一次分裂使得60条anachoreta蛾的染色体分离开来,而curtula蛾的染色体各自分裂,于是各个子细胞分别得到59条染色体;第二次分裂又使59条染色体各自分裂,所得的生殖细胞中会有59条染色体,且都是二倍型。只要继续回交,就有可能产生三倍型个体。虽然在限定条件下,用这种回交的方法,是有可能维持三倍体品系的,但由于这种杂合子的精子在生成过程中有很多不规则性,加之可能导致杂交的后代没有生殖能力,因此,要在自然条件下,建立稳定的三倍体品系,基本上难以实现。
由于基因间的平衡得以保持,我们预计三倍体胚胎的发育应当是正常的。唯一的不和谐因素,可能是三组染色体与遗传所得的细胞质分量两者间的关系。生物自身的调控作用,影响有多深远,我们还不怎么明确,但可以总结如下:至少在植物中,三倍体的细胞要比正常型(二倍体)的细胞大。
还有一些三倍体是源于两个野生型的杂交,其中某一物种的染色体数是另一物种的两倍,这一点将在下一章节中细讲。
[1]三倍体:遗传学名词,是指含有三组染色体的细胞或生物。三倍体生物因难以进行减数分裂形成配子,故常不育。
[2]风信子:天门冬目风信子科风信子属,多年草本球根类植物,鳞茎卵形,有膜质外皮,皮膜颜色与花色呈正相关,未开花时形如大蒜,原产地中海沿岸及小亚细亚一带,喜阳光充足和比较湿润的生长环境,要求排水良好和肥沃的沙壤土等。
[3]均等分裂:有丝分裂中姐妹染色单体或减数分裂中同源染色体对等分开的一种分裂方式。