多倍体蔷薇
从林奈(C. von Linnè,1707—1778)[5]时代开始,分类学家就对蔷薇[6]的分类感到困惑。最近,瑞典植物学家塔克霍尔姆(Täckholm),英国的三位植物学家哈里森(Harrison)和其同事布莱克本(Blackburn),以及蔷薇科专家兼遗传学家赫斯特(Hurst)先后发现,现已确定的若干群蔷薇,尤其是属于canina蔷薇族的那一部分,都是多倍体类型。它们之间的差异,不能完全归于多倍性,其广泛杂交的作用也有一定影响。
最近,塔克霍尔姆对这些蔷薇做了详细研究。首先来看看他的计算。有一种蔷薇,含有最少的染色体数(14条,n=7),我们常将其当作基础型。此外,还存在有着21条染色体的三倍体(3×7),有着28条染色体的四倍体(4×7),有着35条染色体的五倍体(5×7),有着42条染色体的六倍体(6×7),以及有着56条染色体的八倍体(8×7)(如图92)。在成熟分裂期,有一些多倍体会臻于平衡,其所有的染色体都能两两接合成为二价染色体;而在一些有着奇数染色体数量或是偶数染色体数量(假设是杂合子)的多倍体中,只含7条(或14条)二价染色体,其余的染色体在第一次成熟分裂时会保持单染色体的状态。换句话说,当这七类染色体各有4条,6条,8条染色体的时候,染色体间会两两接合,就好像这些类型都是二倍体一样。不管这些染色体源于何处,这些染色体都不会4条,6条,8条地接合在一起。在这些多倍体中,接合在一起的二价染色体会在第一次成熟分裂时分离开来,所得子染色体分别进入两极。在第二次成熟分裂时,每一条子染色体都会分裂为两半,各自进入一极。这些生殖细胞,不管是花粉还是胚珠,都能因此含有半数的原始染色体。因此,如果它们能进行有性繁殖,则其特定的染色体数目可以维持不变。

图92
蔷薇的多倍体序列。
塔克霍尔姆将另一组蔷薇视为杂合子,因为其生殖细胞内发生了种种改变,这表明它们存在不稳定性。其中的一些蔷薇有21条染色体,因此是三倍体。在成熟的早期阶段,花粉母细胞内含有7条二价染色体和7条单染色体。在第一次成熟分裂期,7条二价染色体一分为二,各有7条子染色体分别进入两极;剩余的7条单染色体不再分裂,随机地分布于两极。由此,可得出多种组合方式。从这一点来看,这些组合确实是不稳定的。在第二次成熟分裂期,所有的单染色体,不论是二价染色体分裂而来的子染色体,还是一开始未发生分裂的单价染色体,都中缢成两部分。这样,许多由此得来的子细胞出现退化现象。
还有一些杂合子含28条染色体(4×7),但塔克霍尔姆并未将其划到真正的四倍体种系中,因为接合时期的染色体表明,每类染色体不够4条。该类型只含7条二价染色体和14条单染色体。在第一次成熟分裂期,7条二价染色体分裂开来,剩余的14条单染色体未分裂,且无规律地分布着。
另有一些杂合子含35条染色体(5×7),成熟时有7条二价染色体和21条单染色体(如图93)。这两种染色体的表现,同先前的那个案例一样。
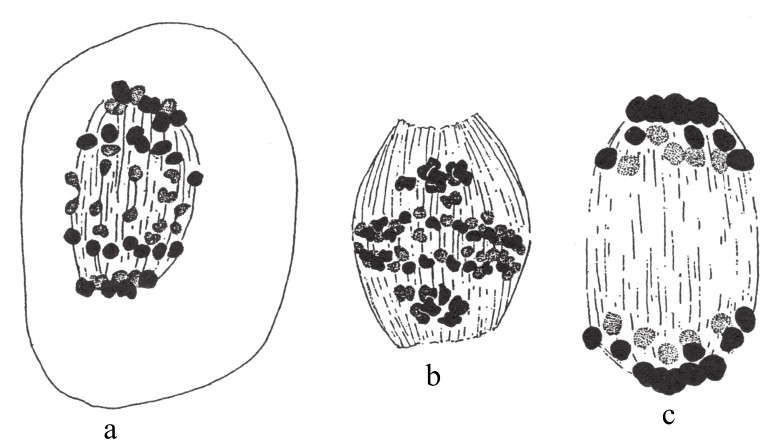
图93
含35条染色体的五倍体蔷薇细胞第一次发生成熟分裂。
第四种杂合子含有42条染色体(6×7),成熟时只有7条二价染色体,而单染色体却有28条。染色体在成熟分裂时期的行为,与先前案例所描述的一样。
现在,从花粉的形成方面,这四种“杂合子蔷薇”可被划分为如下类型:
7条二价染色体+7条单染色体=21条染色体
7条二价染色体+14条单染色体=28条染色体
7条二价染色体+21条单染色体=35条染色体
7条二价染色体+28条单染色体=42条染色体(https://www.daowen.com)
以上这些杂合子的特殊行为,体现在只有14条染色体会接合成7条二价染色体。我们必须假定,这些染色体都是相同的,或者说几乎一样,所以它们能接合到一起。塔克霍尔姆认为,其他各组的7条染色体来自不同野生物种的杂交,否则很难理解这些组的染色体为什么不两两接合。用杂交的方式得来的新增染色体与原来的那一组染色体之间的差异,以及每一组染色体之间的差异,都会阻碍接合的发生。
我们还可以谈一下另外两种杂合子物种,两者都含有14条二价染色体和7条单染色体。在这样的杂合子中,发生接合的二价染色体数是先前那些案例的两倍。
在canina蔷薇群中,只有少量杂合子的胚囊染色体有过历史记载(如图94)。有7条二价染色体位于赤道面上,其余的单染色体(21条单染色体)都集中于一极。当这些二价染色体分离之后,一半去向一极,另一半去向另外一极。结果是,一个子细胞的细胞核含有分裂所得的7条子染色体和所有21条单染色体(共计28条),而另外一个子细胞的细胞核只含7条子染色体。如果这个有着28条染色体的卵细胞继续发育,且与另外一个含有7条染色体的精子(假设这种花粉细胞的精子不发挥作用)结合,那么受精卵会含有35条染色体,即和原始型的染色体数相同。
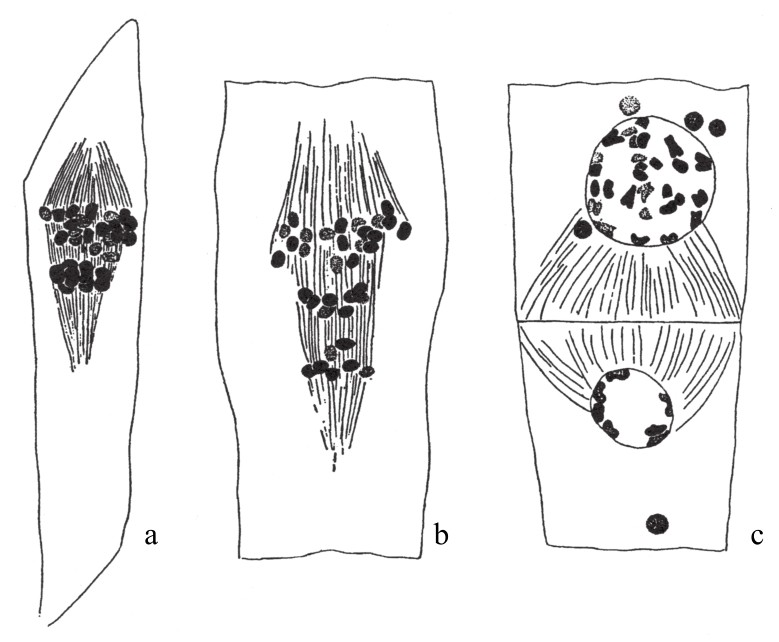
图94
蔷薇卵细胞的成熟分裂。所有的单染色体(21条)都去向一极,这一极还含有接合染色体分裂而来的7条染色体。
这些多倍体杂合子蔷薇的繁殖过程,目前还不能完全阐述清楚。如果通过枝繁殖的方式,那么,它们就能维持在受精过程中得到的染色体数目。通过单性繁殖[7]得来的杂合子种子,也会维持一定数目的体细胞染色体。但当单性繁殖时,花粉和卵细胞在形成时染色体分布的不规则会导致许多不同的组合。如果不知道这些杂合子的染色体之间的相互关系,那么在阐述其遗传过程时,便会遇到困难。即使在这方面的研究有所进展,但对于弄清楚这些杂合子蔷薇的组合方式,仍有很多问题尚待研究。
赫斯特对野生型和栽培型的蔷薇属都有过研究,他认为野生型的二倍体蔷薇属是由五种主系构成[8],可以表示为AA,BB,CC,DD,EE,分别如图95的a-d,e-h,i-l,m-p和q-t。通过五种主系的结合,可以分辨出许多组合。例如,有一种四倍体可以表示为BB和CC,还有一种四倍体可以表示为BB和DD,另外有一种六倍体可以表示为AA、BB和EE,有一种八倍体可以表示为BB、CC、DD和EE。

图95
Canin蔷薇系的五种类型,即a-d,e-h,i-l,m-p和q-t。同一排表示的是同一个品种在花、果实、分枝以及刺和叶的生长处等方面的特征。
赫斯特表示,五种主系中,每个系列都有至少50种可以鉴别的性状。这些性状都可以在杂合子组合中看到。环境因素也可能交替促进一个主系的性状或者另一个主系的性状的形成。赫斯特相信,以这些相互关系为基础,对本属各种进行分类是可能做到的。