其他多倍体序列
除了上面提及的那些多倍体之外,在一些其他种群中,也有关于多倍染色体的变种和物种的研究报告。
我们已知道,山柳菊[9]属有几个品种进行有性生殖,另有几个品种,虽然有时会有正常花粉粒的雄蕊,却是单性生殖。罗森伯格(Rosenberg)在若干产生花粉粒的品种中,对它们的花粉发育情况做了研究。他还检测了这些不同种类杂交所得的杂合子。之后,他还研究了有着18条染色体(n=9)的auricula耳状山柳菊与有着36条染色体(n=18)的aurantiacum山柳菊杂交所得的杂合子的花粉细胞的成熟分裂。在杂合子的第一次成熟分裂期,出现了9条二价染色体和9条单价染色体。但同样有一些杂合子出现例外,或许是由于在祖代aurantiacum山柳菊的花粉细胞中出现了染色体数目异常。在第一次成熟分裂期间,每条二价染色体一分为二,大多数的单染色体也会发生分裂。
罗森伯格对两种四倍体山柳菊(pilosella和aurantiacum,各含36条染色体)杂交所得的子一代杂合子的成熟分裂也做了研究。这些杂合子的体细胞,含38到40条染色体。有两个案例,出现了18条二价染色体和4条单价染色体。在另一组杂交实验中,即有着36条或42条染色体的excellens山柳菊,与有着36条染色体的aurantiacum山柳菊杂交所得的杂合子中,出现了一例含18条二价染色体。所以,极有可能的是其亲代excellens山柳菊含有36条染色体。在另外一组同样的杂交实验中,所得子一代中的花粉大多为不育的,花粉内有大量的二价染色体和很多单价染色体。这样的结果,在其他两个四倍体的杂交中也能观察到。总之,四倍体的杂交结果说明,不同物种中出现的相似染色体更容易彼此接合,或者至少说明,相对于同一物种中的相似染色体来说,二价染色体通过以上途径形成(来自两个物种的相似染色体的接合)是更有可能的。
Archieracium种系中,既存在有性繁殖的物种,又存在单性繁殖的物种,但以单性繁殖较常见。罗森伯格对单性生殖物种的花粉成熟期进行过研究。若是单性繁殖,在其胚囊中不会出现减数分裂,但该细胞仍然保留二倍数目的染色体。花粉在发育过程中,变动很大,且几乎不会出现可生殖花粉。在减数分裂时,花粉母细胞的染色体分布也是紊乱的。罗森伯格对几种无配子生殖[10]的山柳菊物种(其花粉基本上不会有受精能力)的成熟阶段有过研究(如图96)。他解释道,花粉之所以出现这么大的变化(不能发挥受精作用),一方面是由于它们起源于四倍体(大多数类型都会出现二价染色体和单染色体),另一方面是由于所有接合染色体逐步衰退,同时又抑制了一次成熟分裂。据说,卵母细胞也可能出现类似的变化,致使其在单性繁殖时保留所有的染色体数。
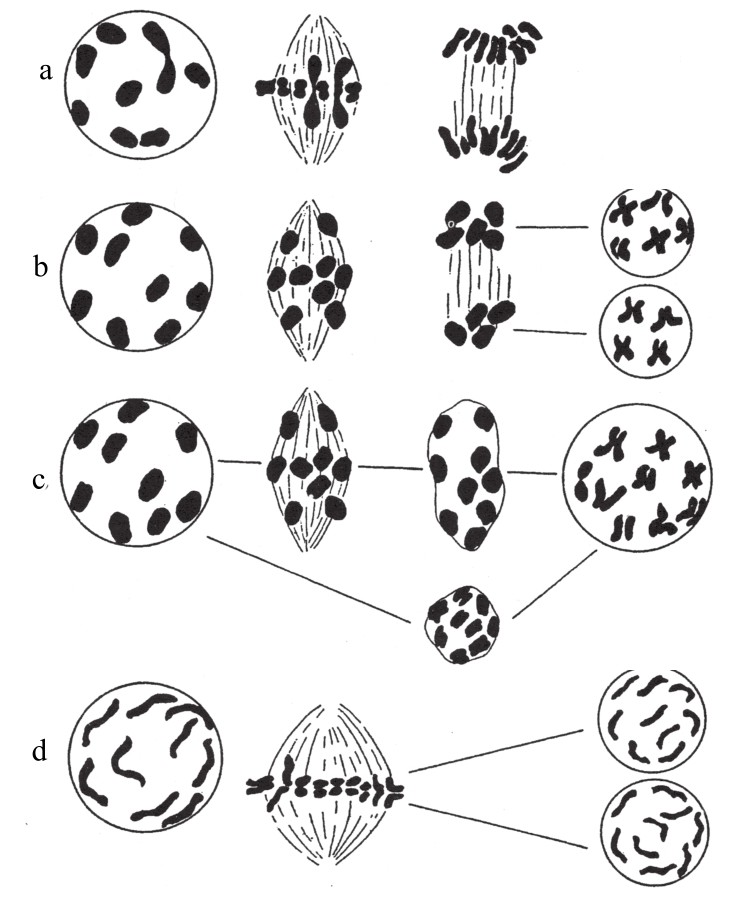
图96
山柳菊几种无受精作用物种的花粉细胞的成熟阶段。
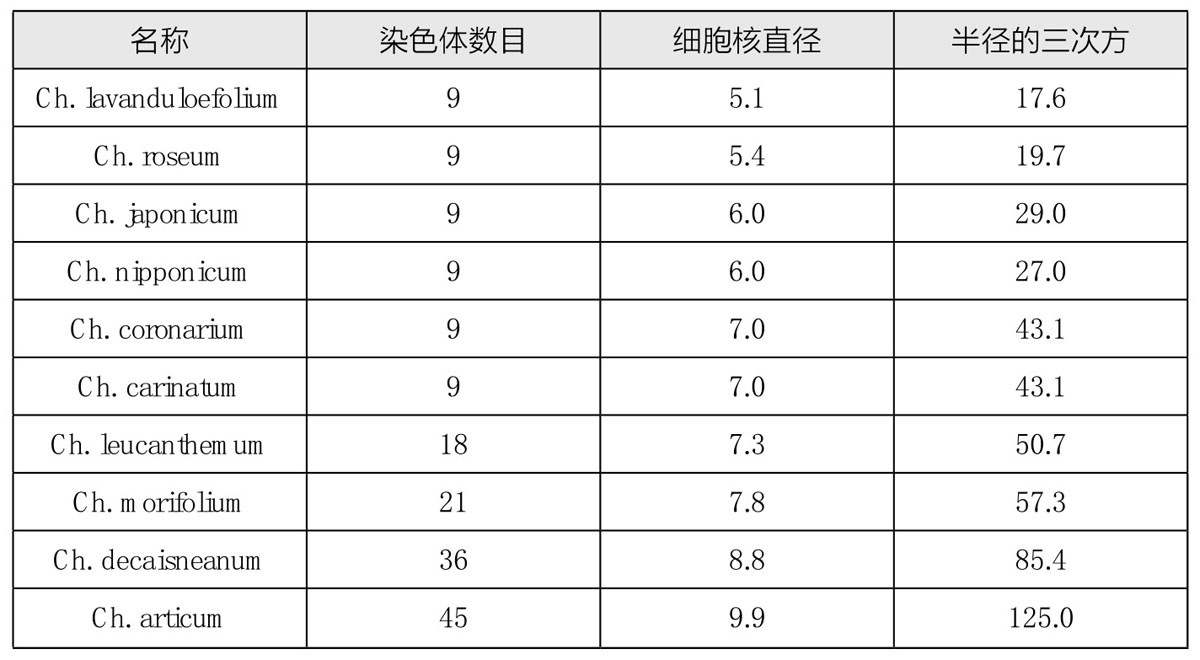
在栽培所得的菊花变种中,田原正人(Tahara)发现了多倍体序列。在这些变种中(如图97),各有9条单染色体,但这9条染色体大小不同。更重要的是,在不同的物种中,染色体的相对体积在这些变种中也是不同的(如图98)。这一点在后面会继续论述。还有就是,在一些有着相同染色体数目的案例中,细胞核的大小却又不同。另有一些菊花,其染色体数是9的倍数(如图99)。其中,有两种是18条染色体,有两种是27条染色体,有一种是36条,有两种是45条。表8为其染色体数目和细胞核大小之间的关系:
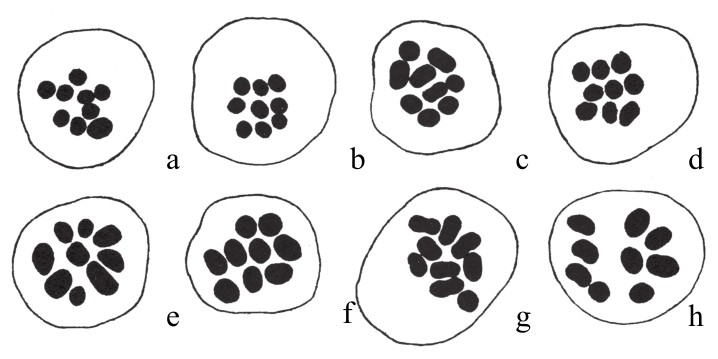
图97
菊花的8种变种,每一种变种都含有减数分裂后的9条染色体。
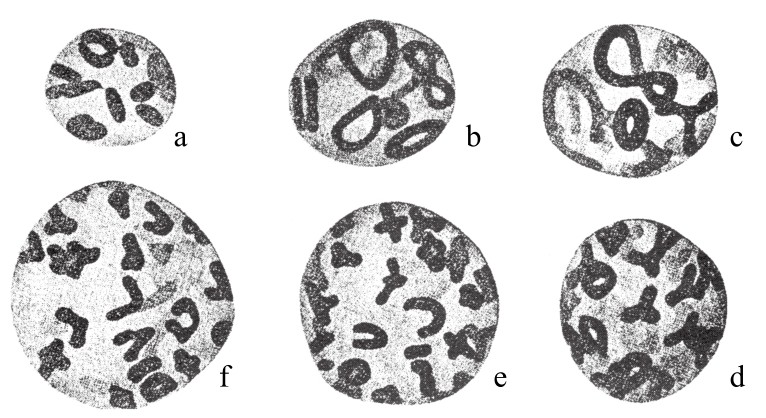
图98
菊花不同变种的多倍染色体群组。图a有9条染色体;图b有9条染色体;图c有18条染色体;图d有21条染色体;图e有36条染色体;图f有45条染色体。
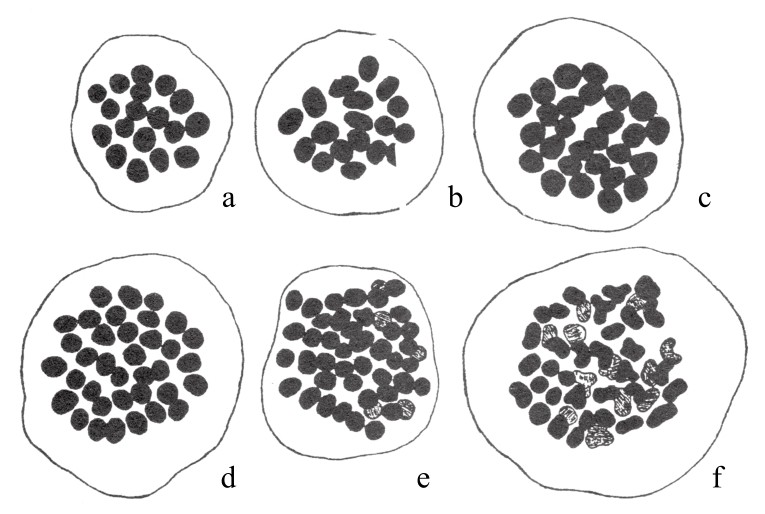
图99
几种菊花变种的终变期细胞核。图a和b有18条染色体,图c有27条染色体,图d有36条染色体,图e有45条染色体,图f有45条染色体。
表8 菊花变种的染色体数目和细胞核体积的关系
大泽一卫(Osawa)曾报告过桑树的三倍体变体。通过对85种桑树变体的研究,他发现有40种都是三倍体。二倍体桑树的染色体数目为28(n=14),三倍体桑树的染色体数目为42(3×14)。二倍体桑树是可育的,然而三倍体桑树在成熟分裂期会表现出不规则性(因为有单价染色体的存在),其花粉粒和胚囊都不能发育成熟。在三倍体的第一次成熟分裂期,不管是花粉细胞还是大孢子母细胞[11],都含有28条二价染色体和14条单价染色体。这14条单价染色体会随机分布于两极,且在第二次减数分裂时缢裂。
在枫叶中,似乎也出现了多倍体。泰勒(Taylor)研究过如下类型的枫叶:有两种含有26条染色体(n=13),有两种含有52条染色体(n=26),还有含144条染色体(n=72),或108条染色体(n=54),或72条染色体(n=36)的品种。泰勒也发现过含其他数目染色体的枫叶品种。
蒂施勒(Tischler)在研究甘蔗时发现,有些品种的单倍数[12]为8条、16条和24条(二价)染色体。据布雷默(Bremer)的研究,另有两种变体,一种含有40条单倍染色体,另一种含有56条单倍染色体。此外,他也报告过含有其他染色体数目的甘蔗品种。在这些组合中,部分或许源于杂交。然而,染色体数目上的差异,究竟有多少是源于杂交,目前我们还不知晓。布雷默还研究过少量杂合子的成熟分裂过程。
据海波恩(Heiborn)所说,薹类carex各物种的染色体数目差异很大,而且没有明显的多倍体序列。“现在最重要的是,要给‘多倍体’这个词下一个清晰明了的定义。从第二章的染色体数目列表中可看出,一系列多倍体有的以3为基数(9,15,24,33,36和42),有的以4为基数(16,24,28,32,36,40和56),有的以7为基数(28,35,42和56),以此类推。但是,作者认为,仅凭这些数字关系,还不能将其视作多倍体的案例。一个多倍体序列的染色体总数,必须是单倍染色体的确切倍数,其由一定组数的相同单倍染色体相加而来。然而,我们都知道,例如pilulifera莎草的染色体并不包含3组(组内相同的)3条染色体,而是由3条长染色体,4条中等染色体和2条短染色体构成;ericetorum莎草也不含这样的5组染色体,而是由1条中等染色体和14条短染色体组成。因此,这两个物种的染色体组并非以增加染色体群的方式来增加染色体,而是以其他方式。”研究人员对于问题较多的其他多倍体序列也有过研究,如酸模属、罂粟属、桔梗属、紫罗兰属、风铃草属和莴苣属。我们发现以下各物种中出现了两种染色体数目,一种是另一种的两倍或三倍:比如车前(6和12),滨藜(9和18),茅膏菜(10和20)和长距兰(21和63)。郎利的最近研究表明,山楂和覆盆子是复杂的多形态物种,有着广泛的多倍性。(https://www.daowen.com)
[1]多倍体:体细胞中含有三个或三个以上染色体组的个体。多倍体在生物界广泛存在,常见于高等植物。多倍体的形成有两种方式,一种是本身出于某种未知的原因而使染色体复制之后,细胞不随之分裂,从而使细胞中的染色体成倍增加,形成同源多倍体;另一种是由不同物种杂交产生的多倍体,称为异源多倍体。同源多倍体是比较少见的。20世纪初,荷兰遗传学家德弗里斯在研究某种待霄草的遗传时,发现一株待霄草的染色体增加了一倍,由原来的24条(2n)变成了48条(4n),成了四倍体植株。它与原来的二倍体植株杂交所产生的三倍体植株是不育的(减数分裂时染色体不配对)。因此这个四倍体植株便是一个新种。
[2]单粒小麦:小麦属中最原始的栽培种,可用作小麦杂交育种的原始材料。
[3]二粒小麦:单粒小麦与一种叫山羊草的杂草杂交的不育后代,由于低温导致其染色体加倍而形成的一个异源多倍体植物。二粒小麦属四倍体种,包括野生二粒小麦和栽培二粒小麦。染色体数2n=4x=28,染色体组型为AABB。
[4]即普通小麦。
[5]林奈:瑞典生物学家,动植物双名命名法的创立者,植物分类学的奠基人。自幼喜爱花卉,曾游历欧洲各国,拜访著名植物学家,搜集大量植物标本。归国后,任乌普萨拉大学教授。
[6]蔷薇:蔷薇属部分植物的通称,主要指蔓藤蔷薇的变种及园艺品种。大多是一类藤状爬篱笆的小花,是原产于中国的落叶灌木,变异性强。
[7]单性繁殖:卵子不经精子的刺激而发育成子代的特殊有性生殖方式,又名孤雌生殖或处女生殖。从发育角度看,卵子不需要受精而发育成新个体(或停顿在胚胎发育的早期)的方式又可称为单性发育。
[8]将这些性状分为五个主要的大类。
[9]山柳菊:多年生草本植物,高30~100厘米。茎直立,单生或少数成簇生,基部常呈淡红紫色,上部伞房花序状或伞房圆锥花序状分枝。
[10]无配子生殖:一种广义的单性生殖,是指维管(束)植物中配子体卵细胞以外的细胞单独分裂并发育产生孢子体的现象。进行减数分裂的无性生殖是指从单倍核的配子体卵细胞以外的细胞发育成形的。以助细胞、反足细胞和原叶体细胞为起源的植物以及蕨类植物的旱蕨属,一般都是以这种方法生殖的。如山柳菊是由受精前的胚乳细胞发育而成的,其生殖过程中配子体可以不经过配子的接合,而直接产生孢子体。
[11]又称为胚囊母细胞。大孢子母细胞是由紧接表皮下的珠心组织的细胞分化而产生的,此细胞比周围的细胞大,其细胞质丰富,可产生核更大的孢原细胞,有的只能成为胚囊母细胞,有的可成为覆盖细胞和胚囊母细胞。
[12]配子细胞核中的染色体数。