2.3.1 大肠杆菌DNA聚合酶Ⅰ及其应用
目前已经从大肠杆菌中分离纯化出三种DNA聚合酶(DNA polymerase,DNA pol),即DNA聚合酶Ⅰ、DNA聚合酶Ⅱ、DNA聚合酶Ⅲ。在这三种DNA聚合酶中,只有DNA聚合酶Ⅰ同分子克隆关系最密切。
大肠杆菌DNA聚合酶Ⅰ是1957年美国的生物化学家A.Kornberg首次从大肠杆菌中发现的,因此又称Kornberg酶。它是由大肠杆菌polA基因编码的一种单链多肽(相对分子质量约为1.09×105),具有三种酶活性。
(1)5'→3'聚合酶活性。当模板为单链DNA及引物(带3'-OH基)或5'端突出的双链DNA,利用体系中的四种脱氧核糖核苷(dNTPs),催化单核苷酸分子逐个加到3'-OH末端,形成3',5'-磷酸二酯键,释放出焦磷酸并使链延长(图2-9)。新链合成的方向为5'→3',与模板DNA的顺序互补。这种聚合作用需要有Mg2+存在。
(2)5'→3'外切酶活性。从5'末端降解双链DNA成单核苷酸或寡核苷酸,每次能切除多达10个核苷酸,这种酶活性在DNA损伤修复中起重要作用(图2-10)。它还可以降解DNA-RNA杂交链中的RNA链,具有RNase H酶活性。这种外切酶活性要求带切除的核酸分子具有游离的5'-磷酸基团,而且核苷酸被切除之前是已经配对的。
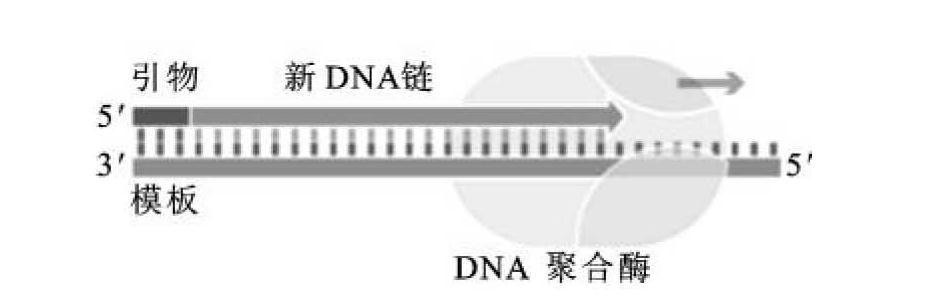
图2-9 大肠杆菌DNA聚合酶Ⅰ的5'→3'聚合酶活性
(3)3'→5'外切酶活性。从游离的3'-OH末端降解单链或双链DNA成单核苷酸。这种外切酶活性在体内DNA复制时主要起校对作用。当DNA复制中掺入的核苷酸与模板不互补而游离时就会被其3'→5'外切酶切除,以便重新在这个位置上聚合对应的核苷酸(图2-11)。这种校对功能保证了DNA复制的真实性,从而降低突变率。该酶切活性虽然对双链DNA也有降解作用,但是在反应体系中有四种dNTP存在,由于聚合活性占有优势,因而阻碍了对双链DNA 3'末端的降解作用。
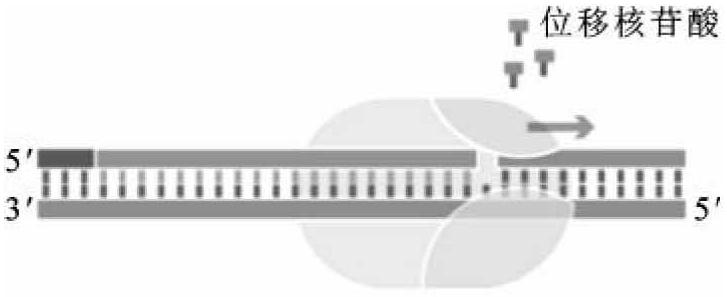
图2-10 大肠杆菌DNA聚合酶Ⅰ的5'→3'外切酶活性
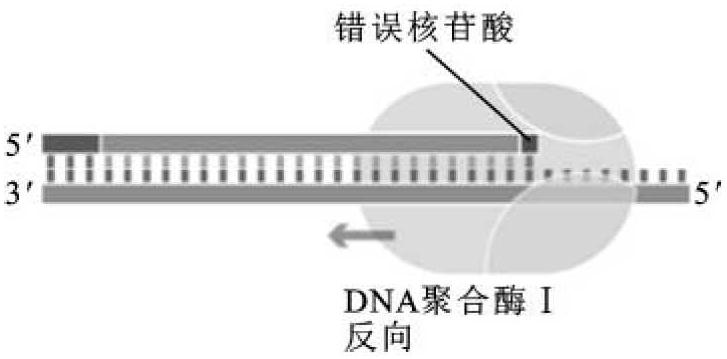
图2-11 大肠杆菌DNA聚合酶Ⅰ的3'→5'外切酶活性
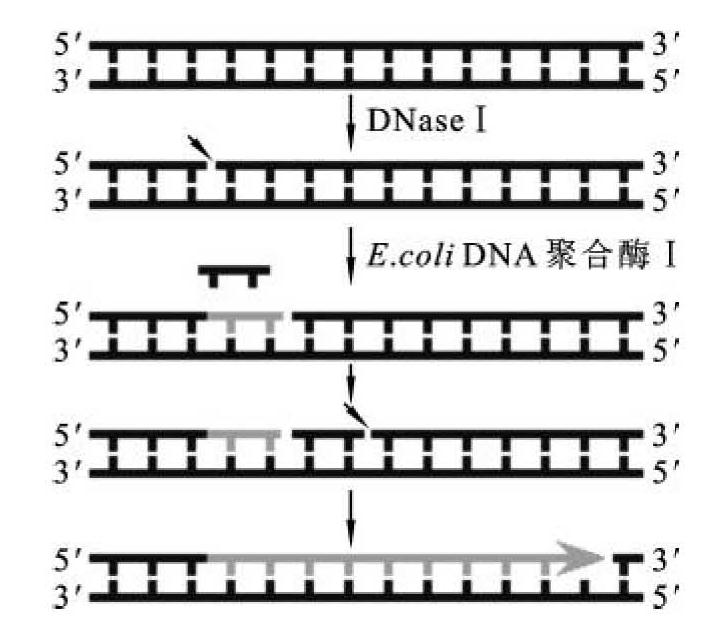
图2-12 大肠杆菌DNA聚合酶Ⅰ的切口平移法示意图
大肠杆菌DNA聚合酶Ⅰ在分子克隆中的主要用途是,通过DNA切口平移,制备供核酸分子杂交用的带放射性标记的DNA探针。在DNA分子的单链缺口上,DNA聚合酶Ⅰ的5'→3'核酸外切酶活性和聚合作用可以同时发生。利用5→3外切活性DNA聚合酶Ⅰ从切口的5'端逐步水解核苷酸时,酶的聚合活性则利用切口的3'游离的羟基逐个加上相应的单核苷酸。但由于DNA聚合酶Ⅰ不能够在3'-OH和5'-P之间形成一个磷酸二酯键,因此随着反应的进行,5'一侧的核苷酸不断地被移去,3'一侧的核苷酸又按序地增补,于是缺口便沿着DNA分子合成的方向移动。这种切口移动的现象特称为切口平移(nick translation)(图2-12)。
应用切口平移法制备DNA杂交探针,其典型的反应体系是,25μL(总体积)中含有1μg纯化的特定的DNA片段,并加入适量的DNaseⅠ、DNA聚合酶Ⅰ、α-32 P-dNTPs和未标记的dNTPs。其中,DNase Ⅰ的作用在于给DNA分子造成断裂或缺口,随后DNA聚合酶Ⅰ则作用于这些单链缺口进行切口平移,使反应混合物中的32P标记的核苷酸取代原有的未标记的核苷酸,并最终形成从头至尾都被标记的DNA分子。这就是所谓的DNA分子杂交探针。
此外,大肠杆菌DNA聚合酶Ⅰ还可用于cDNA第二链的置换合成、3'DNA末端的标记等,但是现在一般用其他酶代替。