3.3.2 λ噬菌体载体
λ噬菌体是迄今被研究得最详细的大肠杆菌双链DNA温和型噬菌体,属长尾噬菌体科。λ噬菌体基因组长度为49 kb,相对分子质量为3.1×107。λ噬菌体的生长周期包括溶源生长和溶菌生长周期(图3-15)。λ噬菌体基因组约含有61个基因,且功能相关基因成簇排列,形成若干个操纵子。基因组两端为cos黏性末端,中间有相当长的DNA片段为溶源生长必需序列,是裂解生长非必需的,因此这段区域为将其改造成外源基因的克隆载体提供了位点。
1.λ噬菌体的形态结构
λ噬菌体基本结构(图3-16)包括头部、包装在头部内的DNA、尾部和尾丝。其中,包围着病毒核酸的头部蛋白质外壳,称为壳体(capsid)或衣壳。壳体由大量同一的壳体蛋白单体分子,即蛋白质亚基(protein subunit)以次级键自组装形成。病毒壳体主要为螺旋对称和二十面体对称两种结构形式,包括二十面体对称的头部和螺旋对称的尾部,尾部连接尾丝。当λ噬菌体感染大肠杆菌时,尾管将基因组DNA注入大肠杆菌,其蛋白质外壳留在细菌细胞外。
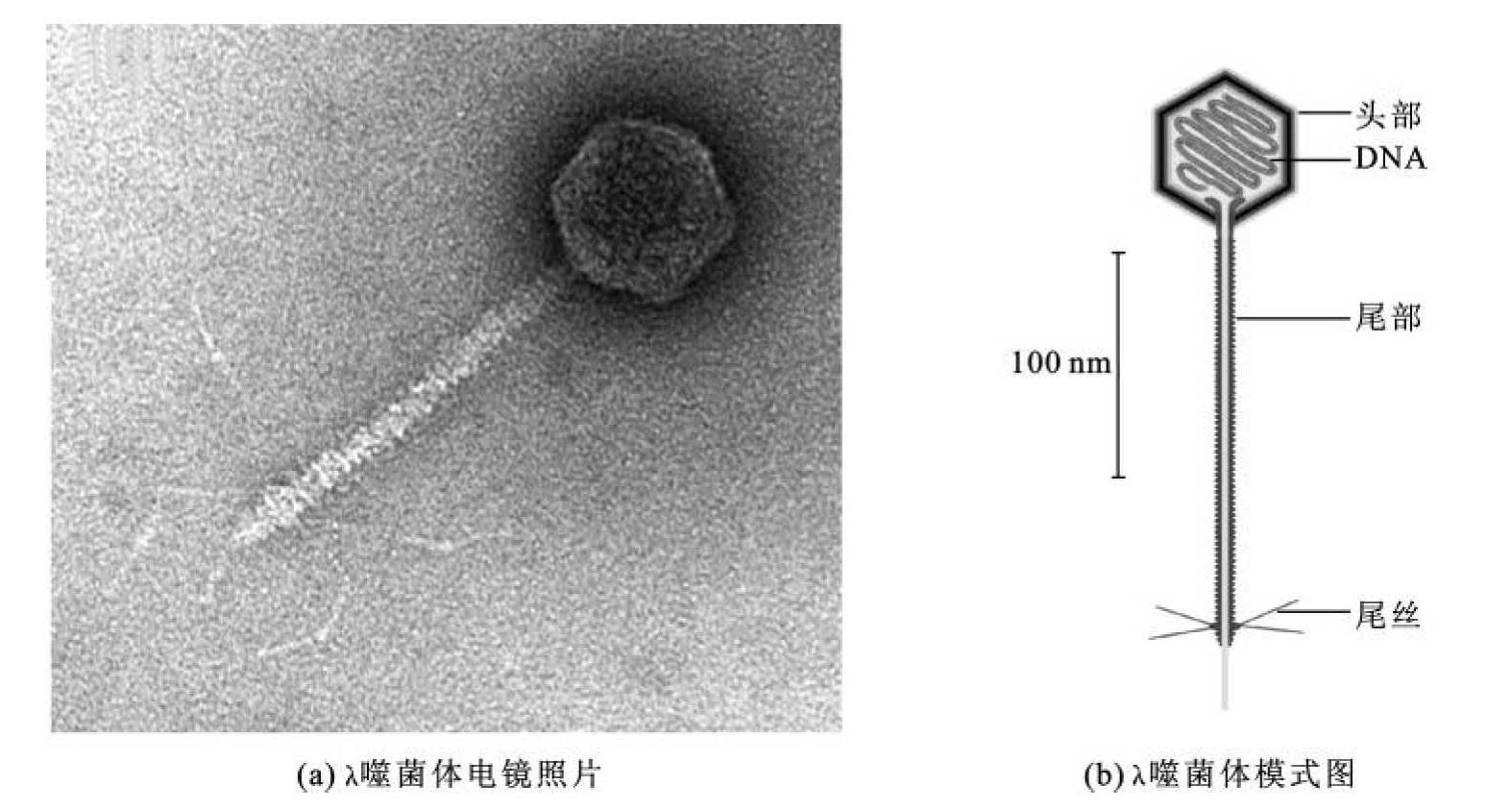
图3-16 λ噬菌体的基本结构
(引自Richard Calendar,2005)
2.λ噬菌体的基因组结构
λ噬菌体的基因组(图3-17)是两端为黏性末端的线状双链DNA,约占噬菌体颗粒质量的54%,相对分子质量为3.1×107,环化状态下为48502 bp,长度为17μm,相似功能的基因聚集成簇,高效利用上游的启动子和操纵子区域。通常将λ噬菌体全基因组分为三部分。
(1)左臂:长约20 kb,从基因A到基因J,编码构成头部、尾部、尾丝等组装成完整噬菌体所需要的蛋白质。
(2)中段:长约20 kb,包含基因J和基因N间的所有基因,编码控制λDNA整合和切出、溶源生长所需的序列,又称非必需区段。
(3)右臂:长约10 kb,位于基因N右侧,是调控区,包含控制溶菌和溶源生长最重要的调控基因和序列,且λ DNA复制起始也在此区域。
左、右臂包含λ DNA复制、噬菌体结构蛋白合成、组装成熟噬菌体、溶菌生长所需全部序列;对溶菌生长来说,中段是非必需的,因此,中段区域是可改造区域,经过改造后成为承载外源DNA的区域。
λ噬菌体基因组的最大特点是在双链线状DNA两端各有12 bp的黏性末端,可以环化,该黏性末端形成的双链区域称为cos位点(cohesive-end site)(图3-18)。λ噬菌体核酸一旦导入细菌细胞,其两端cos位点的黏性末端互补结合,并利用宿主细胞的连接酶将互补区域两端缺口封闭,成为环状DNA分子,进行复制、转录和整合。cos位点黏性末端的可成环结构对λ噬菌体在细菌细胞内复制和溶源生长周期非常重要,如果在体外利用DNA聚合酶将cos位点黏性末端补平成双链,导致其无法成环,DNA就失去其生物活性,将不能进行复制和溶源生长,此结构也是柯斯质粒载体的重要结构基础。
3.λ噬菌体载体的构建
野生型λ噬菌体含有很多优点,使其在分子生物学技术发展的初期就被用于克隆载体,这些优点包括λ噬菌体对大肠杆菌的高效感染性,外壳蛋白、尾部和λDNA的体外自组装性,以及λDNA在大肠杆菌细胞中的高拷贝数复制性等。经过改造的野生型λ噬菌体,消除了其作为载体的限制,同时也增加了其作为克隆载体使用的便利性。
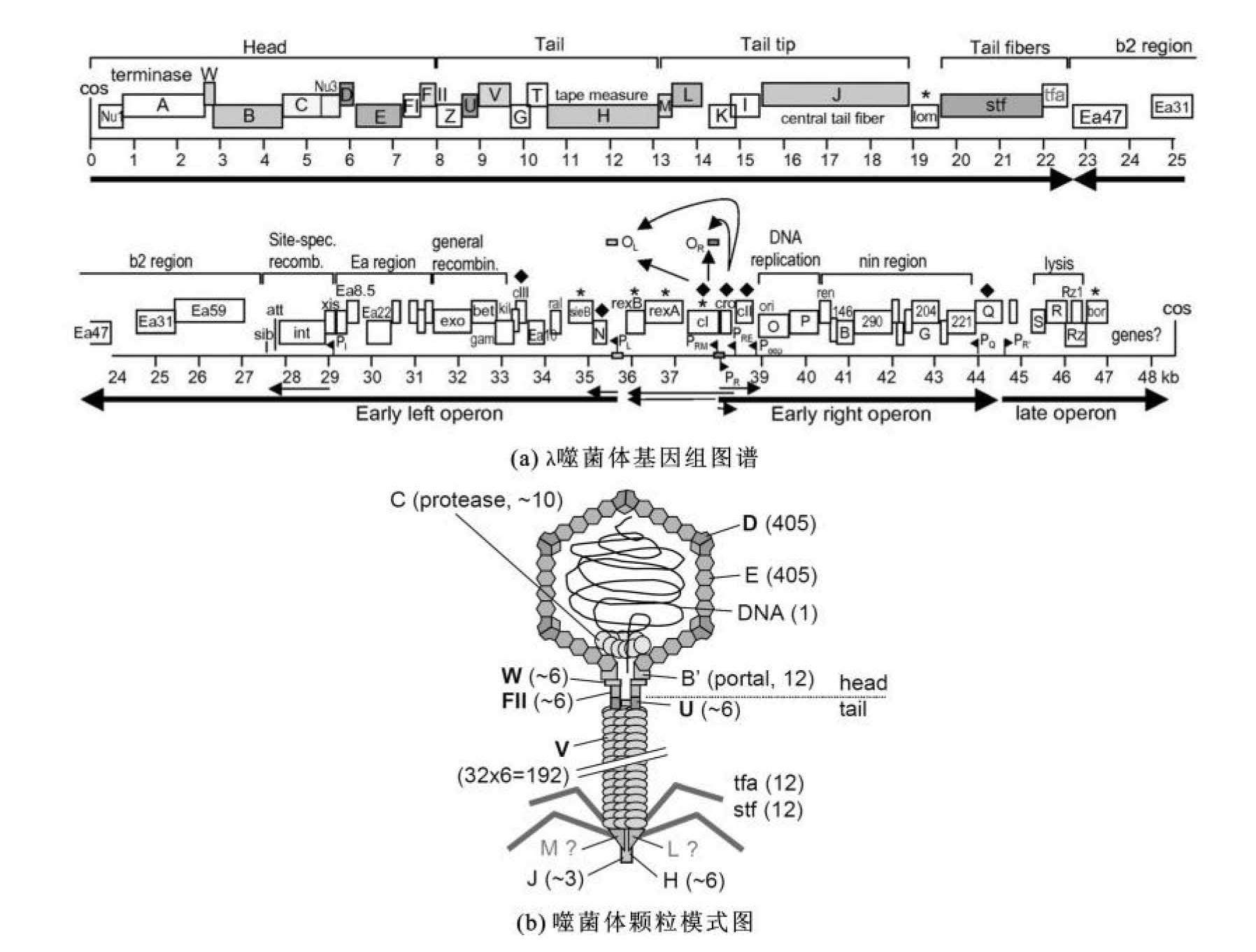
图3-17 λ噬菌体基因组图谱和噬菌体颗粒
(引自Rajagopala S.V.,2011)
(a)灰色标记的开放阅读框(ORFs)编码的相应结构蛋白和(b)图中噬菌体颗粒相应结构的灰度一致。箭头方向为转录本的转录方向。(b)图中展示了组成噬菌体颗粒的各种蛋白质间的相互作用,括号中的数字代表噬菌体颗粒中蛋白质的拷贝数。图中M和L两种结构蛋白目前还不清楚是最终颗粒的组成部分,还是只参与噬菌体颗粒的组装过程。
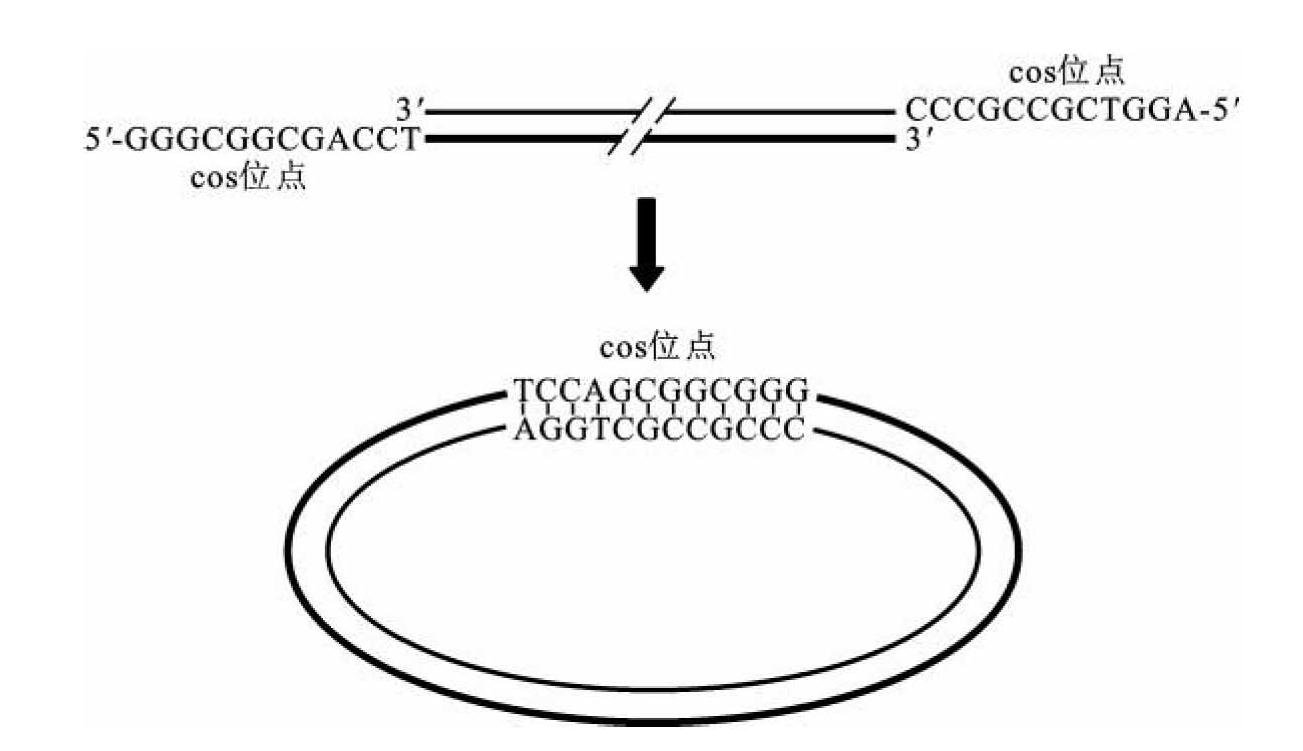
图3-18 λ噬菌体cos位点及其环化过程
λ噬菌体的改造过程主要针对以下几个方面。
(1)删除过多重复的酶切位点,引入多克隆位点(MCS)区域,增加了插入外源DNA的便利性。野生型λ噬菌体的DNA长度,导致其内部单独的酶切位点非常少,其内部还有5个EcoRⅠ和7个Hind Ⅲ,而且个别重复的酶切位点位于噬菌体DNA复制、裂解等重要功能的调控区,因此,为了便于克隆DNA的操作,必须删除多余的酶切位点,或利用点突变的方法消除个别重复的酶切位点。
(2)删除λ噬菌体DNA中段非必需区段DNA,缩小λ DNA长度,以增加外源DNA容纳量。λ DNA的包装范围为正常野生型DNA总量的75%~105%,因此,按野生型λDNA长度为48 kb计算,λ噬菌体的包装上限达到51 kb,因此,野生型的噬菌体作为插入型载体,只能容纳3 kb的外源DNA。λ DNA的改造主要针对中段非必需区段,经过再造后形成两类λ噬菌体载体:
①插入型载体:这类λDNA经过改造后,通常相对分子质量达到噬菌体包装的下限(约38 kb),在这种载体的中段区域只含有一个可供外源DNA插入的位点,外源DNA的重组体需要利用载体上携带的β-半乳糖苷酶α片段基因(lac Z')等选择性标记进行筛选,例如,λgtl0、λgt11、λBV2和λNM540等载体都属于插入型λ噬菌体载体。λNM1149载体含有两个靠近的克隆位点,Hind Ⅲ和EcoRⅠ,也属于插入型载体。此类载体适用于承载10 kb以内的外源DNA,所以主要被用于cDNA和小片段DNA克隆。(https://www.daowen.com)
②替换型载体:这类λDNA经过改造后,大小约40 kb,在这种载体内部含有成对的克隆位点,两个位点间距离大约14 kb,为非必需区段,外源DNA插入载体时会替换两个酶切位点间14 kb λDNA,空载只有26 kb大小,无法进行噬菌体包装,因此重组体无须进行选择型筛选,同时小于10 kb或大于25 kb的重组体也超出了λDNA可包装范围,无法进行λ噬菌体包装。例如,Charon 4、Charon 10和λgtWES等载体属于替换型λ噬菌体载体。替换型载体的最大理论承载量达到24.9 kb,适用于承载较大相对分子质量的DNA片段,因此多被用于克隆真核生物染色体DNA。
(3)引入正向选择标记,便于重组噬菌体筛选,其中常用的选择标记如下:
①β-半乳糖苷酶α片段基因(lac Z')标记基因:来自大肠杆菌的β-半乳糖苷酶基因(lac Z),通过α-互补作用,并配合lac Z缺陷型大肠杆菌受体菌株,可以通过蓝白斑筛选鉴定重组噬菌体。
②cI+标记基因:带有此标记基因的噬菌体配合高频溶源化(hfl-)突变的大肠杆菌菌株使用,非重组噬菌体在hfl-突变菌株内cI+标记基因正常表达,立即建立溶源状态,形成混浊噬菌斑,重组噬菌体的外源DNA插入cI+标记基因内部,导致基因失活,细菌无法进入溶源状态,形成透明噬菌斑。
③Spi-选择法:此筛选和存在于λ DNA非必需区段的red和gam基因有关。这两个基因的表达产物可以抑制λ噬菌体在P2噬菌体溶源性大肠杆菌中生长,即Spi+(sensitive to P2 inhibition)表型。在替换型λDNA载体的使用过程中,λDNA空载体含有red和gam基因,不能在P2噬菌体溶源性大肠杆菌中生长,当外源DNA的替换导致这两个基因缺失后,表型由Spi+转变为Spi-,使重组λ噬菌体可以在P2噬菌体溶源性大肠杆菌中生长。利用这种差异进行重组子克隆的筛选方法称为Spi-选择法。
(4)诱导控制λ噬菌体进入裂解周期的基因突变失活,使噬菌体的溶菌生命周期在实验室可控条件下发生,缩小λ噬菌体的溶菌性宿主范围,减少发生生物污染的概率。将无意突变引入λ DNA控制裂解周期的基因内部,如A、B、E、S和W等基因,配合K12等少数几种具有矫正功能的大肠杆菌,只有在这类宿主细胞中带有突变基因的噬菌体才能繁殖。
4.λ噬菌体载体的包装
外源DNA被连入λ DNA载体后,下一步就是将重组体导入大肠杆菌受体细胞,使λDNA在细胞中复制并包装成成熟的噬菌体。
有两种途径可以将重组后的λ DNA导入宿主细胞:
(1)λ DNA直接导入细胞,这是最为简单的一种途径。这一途径和质粒进入受体细胞的原理一致,受体细胞直接捕获λDNA进入细胞。这种方法的缺点是λ DNA导入受体细胞成功率非常低,由于其相对分子质量比质粒大,因此比质粒的转化效率低。
(2)将重组λ DNA在体外包装成噬菌体颗粒,按照正常噬菌体繁殖过程感染大肠杆菌受体细胞,大大提高λ DNA的转染效率。
λDNA体外包装过程是指噬菌体的DNA、头部、尾部在试管中完成组装,产生成熟噬菌体。噬菌体的头部和尾部的组装是独立完成的,重组λDNA体外包装过程主要是利用了几种基因突变的噬菌体分别合成头部和尾部,再把头部、尾部和λDNA混合孵育完成噬菌体的自组装过程。其中主要涉及两种主要的基因突变噬菌体:
(1)D基因突变的噬菌体:D基因产物存在于λ噬菌体头部的外侧,其功能与头部成熟及λ DNA进入头部有关,其产物占头部总蛋白的20%。D基因突变后,噬菌体组装被阻断在头部成熟的前期,导致λDNA无法进入头部,因此其宿主细胞中积累大量的头部前体蛋白。
(2)E基因突变的噬菌体:E基因的产物是λ噬菌体的头部的主要成分,占头部总蛋白的70%,E基因发生突变后,在其宿主细胞中无法形成头部结构,导致在细胞中积累了大量尾部蛋白。
这两种基因突变噬菌体宿主细胞中的头部前体和尾部恰好是组装为成熟噬菌体的两种必备主要成分。因此,将λDNA和这两种宿主细胞提取出来的头部和尾部混合在一起,在一定的温度条件下便可以组装出成熟的λ噬菌体颗粒。
λ噬菌体DNA包装过程受到λDNA长度的限制,其允许包装的长度范围为野生型λDNA长度的75%~105%。野生型λDNA长度为48 kb,那么包装长度在36~50.4 kb。只有在此范围长度的λDNA包装后的噬菌体颗粒才具有正常活性。