7.1.1 原核生物的转录
1.mRNA转录合成条件
1)底物
mRNA转录是以四种核糖核苷酸(ATP、GTP、CTP和UTP)为底物,通过脱焦磷酸反应,合成mRNA。
2)模板
转录反应一般以一段单链DNA为模板。在原核生物中,这个单链DNA片段可以包括一个乃至几个基因,其中含有一个基因的称为单顺反子,含有多个基因的称为多顺反子。在结构上,基因由多个不同的区域组成,可以划分为编码区和非编码区两个基本组成部分。
(1)编码区:指能转录为相应的mRNA,进而指导蛋白质合成的DNA区域。编码区能够编码蛋白质,由连续的密码子组成,其位置在起始密码子(AUG)和终止密码子(UAA、UAG或UGA)之间。其核苷酸序列根据基因的功能,被称为结构基因。
(2)非编码区:指不能够编码蛋白质的核苷酸序列,非编码区位于编码区的上游及下游。
①位于编码区上游的非编码核苷酸序列。位于编码区上游的非编码核苷酸序列在调控遗传信息表达方面最为重要,它对调控遗传信息的表达是必不可少的。在原核生物中,位于上游的调控序列与一系列受其操纵的结构基因共同形成一个转录单位,称为操纵子。操纵基因是操纵结构基因表达的基因,它是一段能被特异阻遏蛋白识别和结合的DNA序列。特异阻遏蛋白是由调节基因编码,但调节基因不属于操纵子。当特异阻遏蛋白未与操纵基因结合时,由它所控制的结构基因就开始转录、翻译和合成蛋白质。当特异阻遏蛋白与操纵基因结合时,操纵基因处于“关闭”状态,结构基因就停止转录与翻译。启动子是指与RNA聚合酶识别、结合并与起始转录有关的一些DNA序列。以E.coli乳糖操纵子(图7-1)的启动子为例,RNA聚合酶进入位点可以分为两部分,即识别位点和结合位点。其中,Sextama盒(—35序列)为RNA聚合酶的识别位点,序列为TTGACA。Pribnow盒(—10序列)为RNA聚合酶的结合位点,序列为TATAAT。RNA聚合酶一旦与启动子结合,就可以开始催化mRNA转录。此外,在E.coli乳糖操纵子基因的启动子上游还有CAP-cAMP结合位点。CAP(cyclic AMP receptor protein)为环腺苷酸受体蛋白,是一种二聚体蛋白质,亚基相对分子质量为2.3×104。只有在cAMP与此受体结合成的复合物结合到CAP位点后,才能刺激RNA聚合酶与起始部位结合,从而起始转录过程。
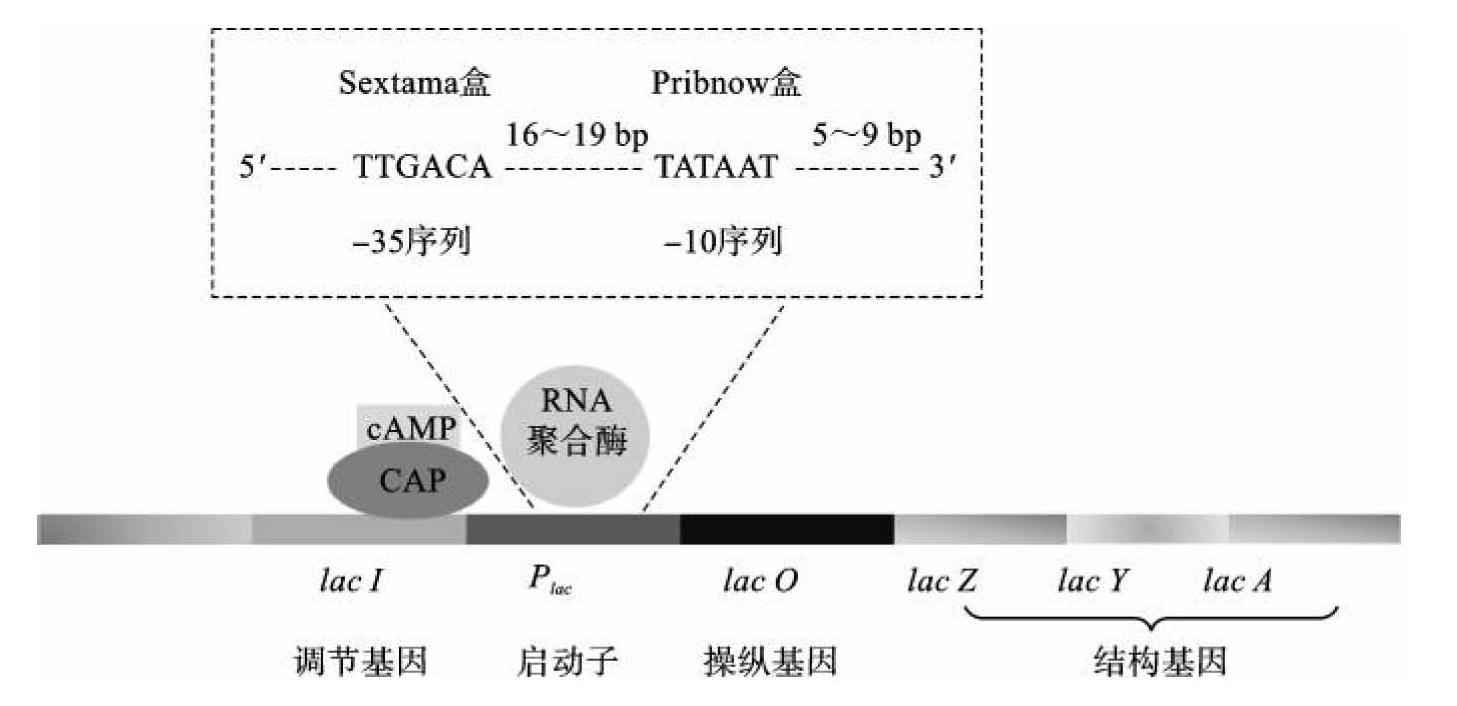
图7-1 E.coli乳糖操纵子结构示意图
②位于编码区下游的非编码核苷酸序列。在一个操纵子的3'末端往往有特定的核苷酸序列,具有终止转录功能,这一序列称为转录终止子,简称终止子。终止子的主要作用是使RNA聚合酶停在DNA模板上不再前进,RNA的延伸也停止在终止信号上,完成转录的RNA从DNA模板和RNA聚合酶上释放出来。对RNA聚合酶起强终止作用的终止子有两个明显的结构特点,即有一段富含AT的区域和一段富含GC的区域。其中,GC富含区域具有回文对称结构,这段终止子转录后形成的RNA具有茎环结构;AT富含区域对应转录出一连串的U。这些结构特征共同促进转录的终止。因此,在构建表达载体时,为了稳定载体系统,防止克隆的外源基因表达干扰载体的稳定性,一般在多克隆位点的下游插入一段很强的rrB核糖体RNA的转录终止子。
3)RNA聚合酶
RNA聚合酶是一种不同于引发酶的依赖于DNA的RNA合成酶。该酶在单链DNA模板以及四种核糖核苷酸存在的条件下,不需要引物,即可按5'→3合成RNA。
大多数原核生物只含有一种RNA聚合酶,它负责几乎所有的mRNA、rRNA和tRNA的合成。原核生物来源的RNA聚合酶的组成是基本相同的。以E.coli为例,RNA聚合酶全酶由6个亚基构成,即σα2ββ'ω,相对分子质量为4.65×105。其中,σ亚基与转录起始位点的识别有关,它的主要功能是确定DNA的模板链和辅助转录的起始,而在转录合成起始后被释放。余下的部分(α2ββ'ω)称为核心酶,负责RNA链的延伸。其中,β亚基和β'亚基共同形成RNA聚合酶的催化中心。α亚基可能与核心酶的组装及启动子的识别有关,并参与RNA聚合酶和部分调节因子的相互作用。
4)终止因子和抗终止因子
(1)终止因子(ρ因子):终止因子是一种由6个相同亚基组成的六聚体蛋白质,其相对分子质量为2.80×105。它具有NTP酶和解螺旋酶活性。ρ因子能水解各种核苷酸三磷酸,它通过催化NTP的水解促使新生RNA链从三元转录复合体中解离出来,从而终止转录。
(2)抗终止因子(N蛋白):Ⅰ噬菌体中由N基因编码的N蛋白具有抗转录终止作用。N蛋白的功能发挥依赖于宿主所产生的NusA、NusB、NusG和S10等几种蛋白质。在终止子附近有一个二重对称序列,N蛋白能识别转录所形成的颈-环结构,并与之结合。NusA以二聚体的形式存在,其一个亚基与RNA聚合酶结合,另一个亚基与N蛋白结合,当其他蛋白质都结合到Nut位点时,便形成一个蛋白质复合物,并通过NusA与RNA聚合酶的结合,改变RNA聚合酶的构象,使之对终止信号不敏感,继续催化RNA链的合成。(https://www.daowen.com)
2.RNA转录合成的特点
RNA转录合成是在RNA聚合酶的催化下,以一段DNA链为模板合成RNA,从而将DNA所携带的遗传信息传递给RNA的过程。其特点如下:
(1)转录的不对称性:指以双链DNA中的一条链为模板进行转录,从而将遗传信息由DNA传递给RNA。其中,作为RNA转录模板合成RNA的那条DNA链称为反义链(模板链),而与之互补的另一条DNA链称为有义链(编码链)。
(2)转录的连续性:RNA转录合成时,以DNA为模板,在RNA聚合酶的催化下,连续合成一段RNA链。转录中不需要RNA引物。合成的RNA中,如只含一个基因的遗传信息,称为单顺反子;如含有几个基因的遗传信息,则称为多顺反子。
(3)转录的单向性:RNA转录合成时,只能向一个方向进行聚合,所依赖的模板DNA链的方向为3'→5',而RNA链的合成方向为5'→3'。
(4)有特定的起始和终止位点:RNA转录合成时,只能以DNA分子中的某一段为模板,故存在特定的起始位点和特定的终止位点,特定的起始位点和特定的终止位点之间的DNA链构成一个转录单位,通常由转录区和相关的调节序列构成。调节序列对基因表达至关重要,因此对转录起始相关的调控序列的了解在设计高效表达载体时是很有意义的。
3.RNA转录合成的基本过程
1)模板识别
模板识别主要是指RNA聚合酶与启动子DNA双链相互作用并与之相结合的过程。原核生物RNA聚合酶中的σ亚基识别转录起始位点—35区的TTGACA序列(Sextama盒),并促使核心酶结合形成全酶复合物。全酶与该区结合后,即滑动至—10区的TATAAT序列(Pribnow盒),并启动转录。
2)转录起始
当RNA聚合酶以全酶的形式与启动子DNA紧密结合后,形成起始复合物,使DNA分子的构象发生变化,促进启动部位的配对碱基解离。当RNA聚合酶进入起始位点后,就可催化四种核苷三磷酸按碱基配对原则互相配对(A-U、C-G、T-A、G-C),分别结合到反义链上。一般新合成的第一个核苷酸往往是腺嘌呤核苷酸,当第二个核苷酸进入DNA模板时,与第一个3'-OH端形成3',5'-磷酸二酯键,并释放出焦磷酸。此时,启动阶段结束,进入延伸阶段。
3)转录延伸
σ亚基脱离复合物,留下的核心酶与DNA的结合变松,因而较容易继续往前移动。核心酶在DNA链上每滑动一个核苷酸距离,即有一个与DNA链碱基互补的核苷三磷酸进入,与前一个核苷酸3'-OH形成3,5-磷酸二酯键。随着转录不断延伸,DNA双链顺次被打开,并接受新来的碱基配对,合成新的磷酸二酯键后,核心酶向前移去,已使用过的模板重新关闭起来,恢复原来的双链结构。核心酶如此不断滑动,RNA链就不断延长。原核生物RNA合成的速度为每秒25~50个核苷酸。核心酶无模板专一性,能转录模板上的任何DNA序列。合成的RNA链对DNA模板具有高度的忠实性。脱离复合物后的σ亚基可与新的核心酶结合,循环地参与起始位点(又称启动子)的辨认作用。
4)转录终止
DNA模板链上有终止信号,在终止信号中有由GC富集区组成的反向重复序列,在转录生成的mRNA中形成相应的发卡结构。此结构可被RNA聚合酶或ρ因子识别,并与之结合,从而阻碍RNA聚合酶的行进,核心酶不再向前滑动,RNA不再延长,转录终止。新合成的RNA、核心酶和ρ因子都从模板DNA上释放出来。核心酶可再与σ亚基结合,又可辨认启动基因合成新的RNA。根据终止子的强弱不同,转录终止可以分为不依赖于ρ因子的终止和依赖于ρ因子的终止。在E.coli中,ρ因子依赖型终止子占所有终止子的一半左右。