3.2.1 早期对天然质粒的研究
对基因工程载体的研究始于1946年J.Lederberg对大肠杆菌K12细胞中F因子的研究。随着研究的深入,在细胞中发现了多种可以控制细菌特定性状的遗传因子,但当时对这种游离于细菌基因组之外的遗传因子的命名并不统一。直到1952年,J.Lederberg建议将游离于染色体之外的所有遗传因子统一称为“plasmid”,即质粒。目前所发现的众多天然质粒中,人们了解比较详细的主要有以下三类。
1.F质粒
F质粒即F因子或性因子(sexual plasmid),其编码的基因可以调控细菌细胞通过接合作用将含有F质粒细胞中的遗传物质和F因子以较高的效率转移到不含F因子的细菌细胞中,细菌人工染色体(BAC)也是基于F质粒构建的。
F质粒控制细菌细胞性菌毛的形成(图3-1)。性菌毛比普通菌毛粗而长,中空呈管状,数量少,一个细胞仅具1~4根。性菌毛是细菌传递游离基因的器官,作为细菌接合时遗传物质的通道。一般用纤毛表示普通菌毛,而菌毛多指性菌毛。
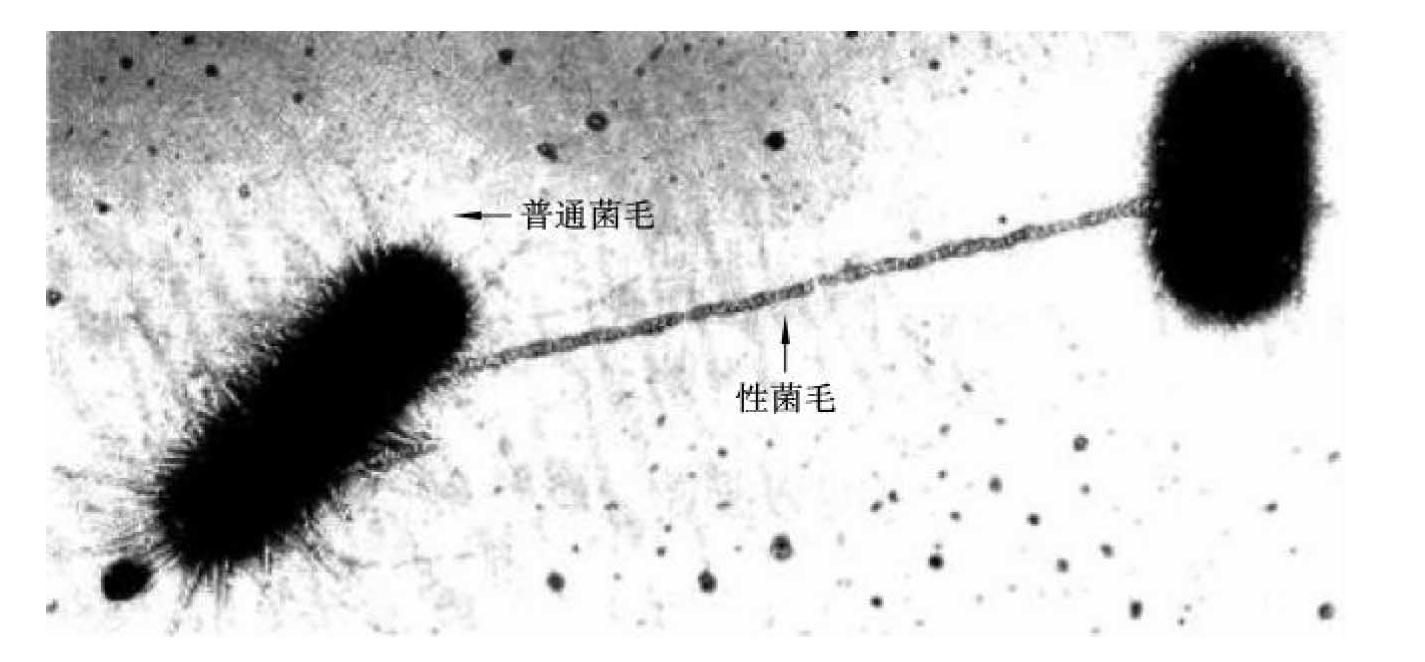
图3-1 F质粒控制细菌性菌毛的形成
带有性菌毛的细菌称为F+菌或雄性菌,无性菌毛的细菌称为F-菌或雌性菌。F+菌体内的质粒或染色体DNA可通过中空的性菌毛进入F-菌体内,这个过程称为接合。细菌的毒性及耐药性等性状可通过此方式传递,这是某些肠道杆菌容易产生耐药性的原因之一。其功能有的是作为两菌接合时传递遗传物质的通道,有的是作为某些噬菌体感染宿主菌的受体。(https://www.daowen.com)
2.R质粒
R质粒是抗药性质粒(耐药质粒)的统称。1975年日本学者Nakaya等首先在福氏志贺菌26(222/CTS)菌株中分离到R质粒,并发现R质粒可以通过细菌接合作用将R质粒上编码的抗生素抗性基因转移到其他敏感菌,从而使其他细菌细胞获得某种抗生素的抗药性。R质粒编码一种或多种抗生素抗性基因,能使获得R质粒的细菌细胞产生对特定抗生素的抗性。这也是众多基因工程载体利用抗生素进行筛选的基本原理。
R质粒分为接合型和非接合型两种类型。接合型R质粒由两部分组成:第一部分是与接合及DNA转移相关的基因,这组基因与F质粒中tra基因十分相似,称为抗性转移因子(resistance transfer factor,简称RTF);第二部分为抗性基因,称为抗性决定因子(R-determinant,简称RD)。不同接合型R质粒中的RTF不同,但具有很高的同源性,而RD则更加多样化。RD的侧翼序列可以介导RD在不同R质粒间转移,有些R质粒甚至可以同时具有两种不同的RD,形成复合R质粒。复合R质粒对多种抗生素和药物具有抗性。由于RD结构具有在不同R质粒间的转移性,许多R质粒甚至可以在异种或异属间转移,因此在一定选择压力下R质粒的抗性可以快速进化。病原菌抗性的进化对人类健康构成很大威胁。
3.Col质粒
Col质粒是一种常见的天然非接合型质粒,其相对分子质量比F质粒和R质粒小,编码大肠杆菌素蛋白。这种细菌毒素蛋白可以杀死和Col质粒宿主亲缘关系密切的不含Col质粒的细胞。