9.4.1 转化植物细胞的筛选
植物外植体经过遗传转化后,只有极少数被转化,这就需要采用特定的方式将转化细胞与非转化细胞区分开来,淘汰非转化细胞。目前,转化细胞与非转化细胞的区分及非转化细胞的淘汰常采用抗生素抗性基因及除草剂抗性基因等,总称选择标记基因(selectable marker gene)。选择标记基因是指可使转化细胞获得其亲本细胞所不具备的新的遗传特性,从而使得能利用特定的选择培养基将转化细胞从亲本细胞群体中选择出来的一类特殊的基因。
1.常用选择标记基因的种类
1)抗生素抗性基因
这类选择标记基因是通过编码一种或多种酶来解除抗生素的毒性,使得已经转入了此选择标记基因的细胞,在有抗生素存在的情况下能够存活。
抗生素抗性基因和其对应的抗生素构成一个筛选体系。使用最多的抗生素抗性基因有npt Ⅱ基因(产生新霉素磷酸转移酶,抗卡那霉素、G418、巴龙霉素、新霉素)、hpt基因(产生潮霉素磷酸转移酶,抗潮霉素)、cat基因(产生氯霉素乙酰转移酶,抗氯霉素)、spt基因(产生链霉素磷酸转移酶,抗链霉素)和gent基因(抗庆大霉素)等。
新霉素磷酸转移酶基因(npt Ⅱ)与G418筛选体系:G418属于氨基糖苷类抗生素,该类抗生素能干扰植物细胞叶绿体及线粒体中蛋白质的合成,从而影响植物的光合作用和能量代谢,导致植物细胞死亡。nptⅡ编码的产物能够修饰该类抗生素分子,从而影响该类抗生素功能的发挥而使抗生素失活。所以在含有G418的培养基上,只有那些转化成功的细胞能够存活下来,而没有转化成功的细胞则因能量代谢受到影响而死亡。
新霉素磷酸转移酶基因在植物遗传转化中应用最早,Bevan M.W.等在1983年已将其用作植物遗传转化的标记基因。美国食品和药物管理局(FDA)在1994年批准的首例商业化应用的转基因延熟番茄就是用该基因作为标记基因。目前,该基因仍然是最常用的转基因植物的选择标记基因。
潮霉素磷酸转移酶基因(hpt)和潮霉素B筛选体系:编码的潮霉素磷酸转移酶(HPT)能够解除抗生素潮霉素B的毒性而起到选择标记的作用。潮霉素B在植物基因工程中的应用有对愈伤组织的选择效果明显,并且对植株的分化影响小等优点。潮霉素磷酸转移酶和潮霉素B在植物基因工程中已经发展成为一个非常有效、作用广泛的筛选体系,在单子叶植物、双子叶植物和裸子植物中均有应用。
2)除草剂抗性基因
这类基因能够赋予转化成功的个体抗除草剂的性状。相比于抗生素抗性的筛选体系,除草剂抗性的筛选体系在转基因植株后代纯化筛选中有其独到的优势。近年来常用的除草剂抗性基因有epsps基因、bar基因和gox基因等。
epsps基因和草甘膦筛选体系:epsps基因编码的酶称为5-烯醇式丙酮酸莽草酸-3-磷酸合成酶,该酶在植物自身氨基酸合成过程中发挥重要作用,该酶的缺失会造成植物因氨基酸的缺乏而死亡。草甘膦是目前世界上使用最广泛的除草剂之一,它能特异性地抑制植物中该酶的活性,阻断氨基酸的合成,造成氨基酸的缺乏,从而导致植株死亡。但将能够过量表达该酶的突变基因转入植物以后,在一定的草甘膦浓度下,会有一部分细胞因该酶不会被抑制而能正常合成氨基酸,并正常生长。
bar基因和草丁膦筛选体系是目前应用最广泛的除草剂类选择标记筛选体系,尤其是对于禾谷类粮食作物特别有效。它编码的膦丝菌素乙酸转移酶,通过乙酸化作用使膦丝菌素失去毒性。表达bar基因的植物转化细胞,在经受致死剂量的膦丝菌素处理之后,仍能正常生长或是以接近正常的速度生长,而敏感的非转化植株则迅速停止生长,并在10~21 d内死亡。特别注意的是要避免产生抗除草剂的转基因杂草。
2.常用选择标记基因潜在的危险性
随着转基因产品的商品化与市场化,转基因农作物的安全问题日益受到人们的关注,利用选择标记基因得到所需要的转化植株之后,选择标记基因成为多余的,甚至是有害的。选择标记基因的潜在危险性主要包括以下几个方面。
(1)可能转移、扩散。例如:抗生素抗性基因在环境中的传播是否影响抗生素治疗的有效性;除草剂抗性标记基因通过花粉和种子等途径在种群之间扩散,可能转移到杂草,产生抗除草剂的“超级杂草”;或者向其他植物中转移,从而对生态环境和生物多样性产生潜在的危害。尽管风险评估报告对一些选择标记基因提供了安全保证,但还是不能彻底消除人们对转基因植物的担忧,而且对选择标记基因及其产物的风险评估是一个既昂贵又烦琐的过程。
(2)影响转化细胞再生。选择标记基因同目的基因共同转化,然后将转化后的细胞培养在含有抗生素或除草剂的培养基上,非转化细胞在逐渐凋亡过程中能够分泌毒素或生长抑制剂,阻碍营养物质向转化细胞的运输,进而影响转化细胞的增殖及分化。
(3)影响多重转化。在细胞第一次转化中使用了某一特殊的选择标记基因,在随后的转化中就要使用不同的选择标记基因,这就给多基因转化造成困难。另外,通过含不同外源基因的转基因植株杂交,可以获得具有多个优良性状的转基因植株,同时植株中选择标记基因的拷贝数也随之增加,多个同源基因的堆积会大大增加基因沉默的可能性。
3.解决选择标记基因安全性的策略
选择标记基因的安全性是近年来国际上关注的热点,当前解决选择标记基因安全性的主要策略有以下两种。
1)选择标记基因的消除(https://www.daowen.com)
从转基因植物中消除选择标记基因的方法有共转化、位点特异性重组、转座子介导的再定位和染色体内重组等。
(1)共转化(co-transformation):将选择标记基因和目的基因分别构建在不同载体或同一载体的不同T-DNA区域,共同转化受体细胞,通过筛选和分子鉴定获得共整合植株。如果这两个基因整合到不同的位点,通过杂交能使目的基因与选择标记基因分离开来(图9-15)。这样,无选择标记的转基因植株可以在子代水平上分离出来。农杆菌介导的转化方法比基因枪转化方法更能有效地将目的基因与选择标记基因分离。
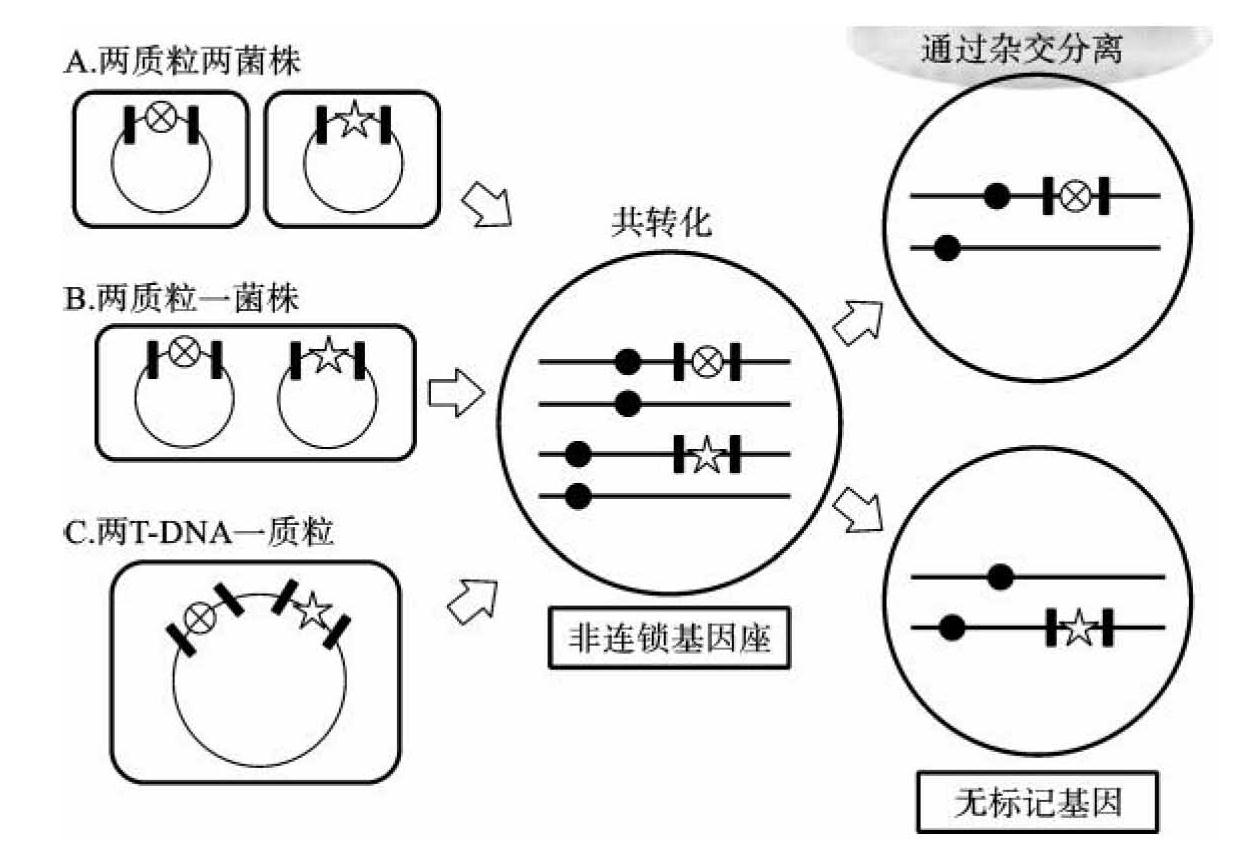
图9-15 共转化消除选择标记基因
(引自Komamine)
(2)位点特异性重组系统(site-specific recombination system):该系统由重组酶及其识别位点组成。同源重组是双向的,通过重组酶作用在DNA专一性位点间实现同源交换,可以把外源基因整合到染色体上,也可以从染色体上把外源基因切除。目前应用于植物遗传转化的重组酶系统有大肠杆菌噬菌体P1的Cre-lox系统、酿酒酵母2 μm质粒的FLP-FRT系统、接合酵母pSR1质粒的R-Rs系统、链霉菌属噬菌体ΦC31的位点特异性重组系统和Mu噬菌体的Gin重组系统。
噬菌体P1的Cre-lox系统是一个位点特异性重组系统,它有两个组成部分:重组酶(Cre)及其识别位点(lox P)。Cre介导重组事件的发生并引起位于两个相邻lox P位点之间的DNA片段的切除。将一个选择标记基因插入两个相邻lox P位点之间,再与一个目的基因相连,导入植物,获得转基因植株。通过异花授粉或再转化将cre基因导入转基因植株,以切除选择标记基因。然后再通过杂交将cre基因与目的基因分离开(图9-16)。
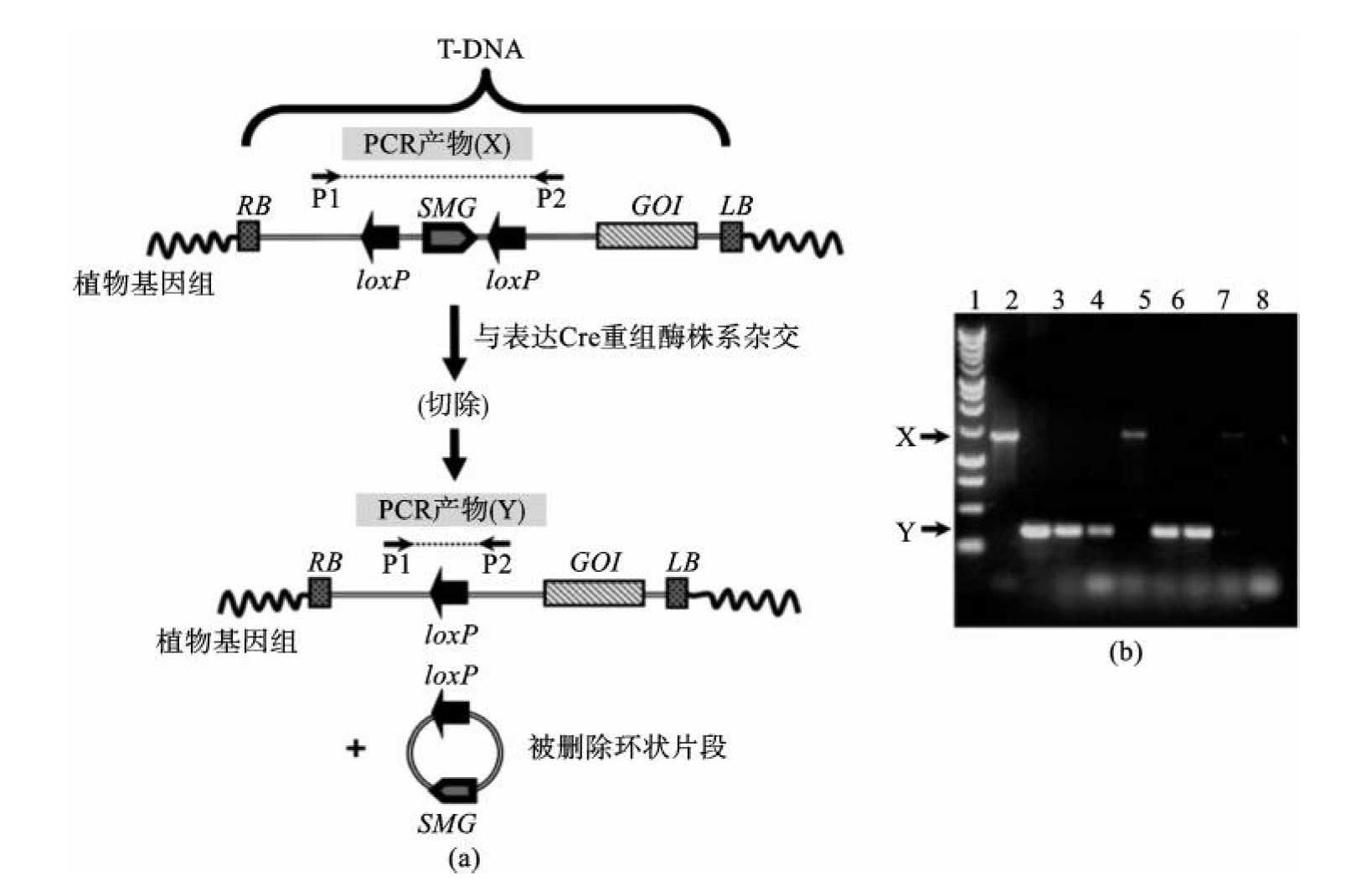
图9-16 位点特异性重组消除选择标记基因
(引自Yuan-Yeu Yau)
GOI:目的基因;SMG:选择标记基因
(3)转座子介导的再定位(transposon-mediated repositioning):该方法是借助于转座子系统如玉米的Ac-Ds、Spm-dSPm的转座作用。非自主的Ds元件是玉米转座元件Ac-Ds家族中的一员,只有在自主的Ac元件存在下,Ds才能在基因组内从所在位点上切除下来并转移到其他位点上。
Yoder等应用Ac-Ds转化系统在相连的两个基因整合后将目的基因或选择标记基因转移到新的位点上,如果新位点距离原位点足够远,通过杂交能够将目的基因与选择标记基因分离开。无选择标记的转基因植株在子代水平上分离出(图9-17)。
(4)染色体内重组系统:在没有外源重组酶表达的情况下,只要两条DNA序列相同或相近,重组就可以在此序列中的任何一点发生。把标记基因放在两个DNA同源序列之间,发生同源重组后,标记基因即被去除,这样的细胞经过诱导再生植株,可得到无选择标记转基因植物。来源于噬菌体的重组结合位点称为attP。通过两个attP的正向重复,将选择标记基因设计在两个attP序列之间,通过attP同源序列介导的染色体内同源重组,实现对选择标记基因的剔除。植物细胞核中同源重组发生的频率很低,在转基因植物的组织培养时期利用染色体内重组系统切除核基因组中的标记基因,需要经过两轮连续的筛选,且效率不足1%。但叶绿体中同源重组频率比较高,因此这种方法在叶绿体转基因去除标记基因方面有很大的应用潜力。
2)利用无争议的生物安全标记基因
目前发现的生物安全标记基因主要有绿色荧光蛋白基因、甜菜碱醛脱氢酶基因、6-磷酸甘露糖异构酶基因、木糖异构酶基因、核糖醇操纵子、谷氨酸-1-半醛转氨酶基因、异戊烯基转移酶基因和吲哚-3-乙酰胺水解酶基因等。
(1)甜菜碱醛脱氢酶(betaine aldehyde dehydrogenase,BADH)基因:它是植物本身就具有的基因,能催化有毒的甜菜碱醛转变成无毒的甘氨酸甜菜碱,作为一种渗透保护剂,还可以提高转基因植株的再生率。
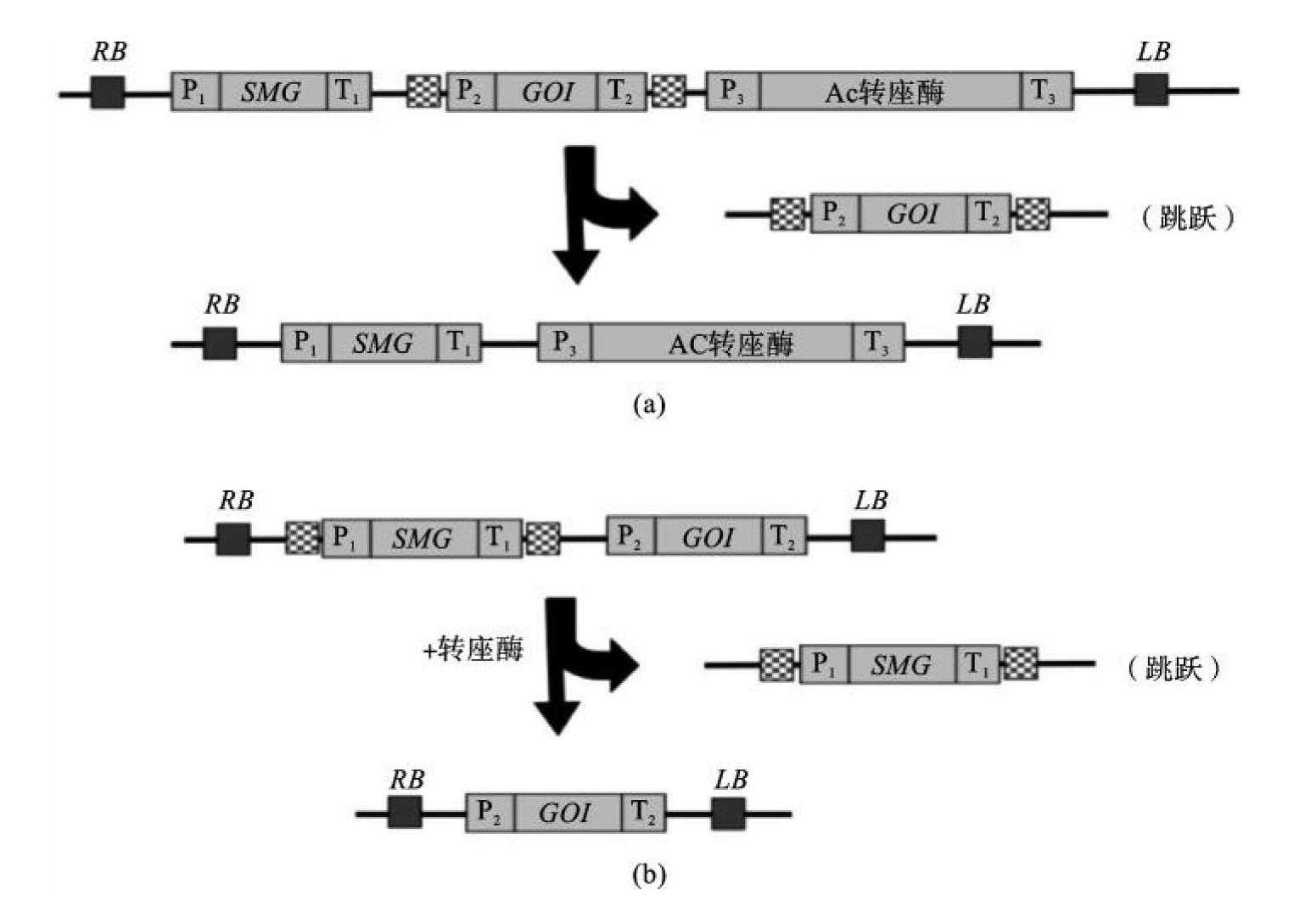
图9-17 Ac转座子消除选择标记基因
(引自Yuan-Yeu Yau)
(2)与糖代谢途径相关的基因:离体培养的细胞不能进行光合作用,必须在培养基中添加一定浓度的碳源(如蔗糖、麦芽糖、葡萄糖等)后细胞才能进行正常的生长分化。近年来,利用这一点研究开发了三种非抗生素标记基因,即6-磷酸甘露糖异构酶(6-phosphomannose isomerase)基因(pmi)、木糖异构酶(xylose isomerase)基因(xflA)和核糖醇操纵子(ribitol operon),它们分别能使转化细胞利用6-磷酸甘露糖、木糖和核糖醇为碳源正常生长,而非转化细胞由于不含有这些基因,不能利用这些碳源,会产生碳饥饿而不能正常生长,从而达到高效选择的目的。
(3)与激素代谢途径相关的基因:包括异戊烯基转移酶(isopentenyl transferase,IPT)基因(ipt)、吲哚-3-乙酰胺水解酶(indole-3-acetamide hydrolyse,IAAH)基因(iaaH)等。ipt基因从农杆菌的T-DNA中克隆而来,编码IPT,参与植物细胞分裂素的合成。细胞分裂素可以促进器官发生,转化细胞在未加细胞分裂素的培养基上能继续生长,形成不定芽。相反,非转化细胞在不含细胞分裂素的培养基中不能正常生长和分化而死亡。IAAH也是通过调节植物体内激素代谢而使转化细胞与非转化细胞的生长与分化造成一定的差异,从而达到筛选出转化细胞的目的。