6.1.2 转化
重组DNA分子在体外构建完成后,必须导入特定的受体细胞,使之无性繁殖并高效表达外源基因或直接改变其遗传性状,这个导入过程及操作统称为重组DNA分子的转化(transformation)。
1.转化的基本概念
重组DNA技术中的转化是指将重组DNA分子人工导入受体细胞的操作过程,它沿用了自然界细菌转化的概念,但无论在原理还是在方式上均与细菌自然转化有所不同,同时也与哺乳动物正常细胞突变为癌细胞的细胞转化概念有着本质的区别。重组DNA人工导入受体细胞有许多方法,如受体细胞的电穿孔和显微注射等,这些导入方法在重组DNA技术中统称为转化操作。
经典的细菌转化现象是1928年英国的细菌学家Griffich在肺炎双球菌中发现的,并在1944年由美国的Avery形成完整的转化概念。细菌转化的本质是受体菌直接吸收来自供体菌的游离DNA片段,并在细胞中通过遗传交换将之组合到自身的基因组中,从而获得供体菌的相应遗传性状,其中来自供体菌的游离DNA片段称为转化因子。具有转化能力的DNA片段常常是双链DNA分子,单链DNA分子很难甚至根本不能转化受体菌。就受体菌而言,只有当其处于感受态(受体细胞最易接受外源DNA片段而实现转化的一种特殊生理状态)时才能有效地接受转化因子。处于感受态的受体菌,其吸收转化因子的能力为一般细菌生理状态的1000倍以上,而且不同细菌间的感受态差异往往受自身的遗传特性、菌龄、生理培养条件等诸多因素的影响。
细菌转化的全过程(图6-1)包括:①感受态的形成。典型的革兰氏阳性菌由于细胞壁较厚,形成感受态时细胞表面发生明显的变化,出现各种蛋白质和酶类,负责转化因子的结合、切割及加工。感受态细胞能分泌一种小相对分子质量的激活蛋白或感受因子,其功能是与细胞表面受体结合,诱导某些与感受态有关的特征性蛋白质(如溶菌酶)的合成,使细菌胞壁部分溶解,局部暴露出细胞膜上的DNA结合蛋白和核酸酶等。②转化因子的结合。受体菌细胞膜上的DNA结合蛋白可与转化因子的双链DNA结构特异性结合,单链DNA或RNA、双链RNA以及DNA-RNA杂合双链都不能结合在膜上。③转化因子的吸收。双链DNA分子与结合蛋白作用后,激活邻近的核酸酶,一条链被降解,而另一条链则被吸收到受体菌中,这个吸收过程为EDTA所抑制,可能是因为核酸酶活性需要二价阳离子的存在。④整合复合物前体的形成。进入受体细胞的单链DNA与另一种游离的蛋白因子结合,形成整合复合物前体结构,它能有效地保护单链DNA免受各种胞内核酸酶的降解,并将其引导至受体菌染色体DNA处。⑤转化因子单链DNA的整合。供体单链DNA片段通过同源重组,置换受体染色体DNA的同源区域,形成异源杂合双链DNA结构。革兰氏阴性菌细胞表面的结构和组成均与革兰氏阳性菌有所不同,供体DNA进入受体细胞的转化机制还不十分清楚。革兰氏阴性菌在感受态的建立过程中伴随着几种膜蛋白的表达,它们负责识别和吸收外源DNA片段。研究表明,嗜血杆菌和奈氏杆菌均能识别自身的DNA,如嗜血杆菌所吸收的自身DNA片段中都有一段11 bp的保守序列5'-AAGTGCGGTCA-3'。这表明革兰氏阴性菌在转化过程中对供体DNA的吸收具有一定的序列特异性,受体细胞只吸收自己或与其亲缘关系很近的DNA片段,外源DNA片段可以结合在感受态细胞的表面,但极少能吸收。
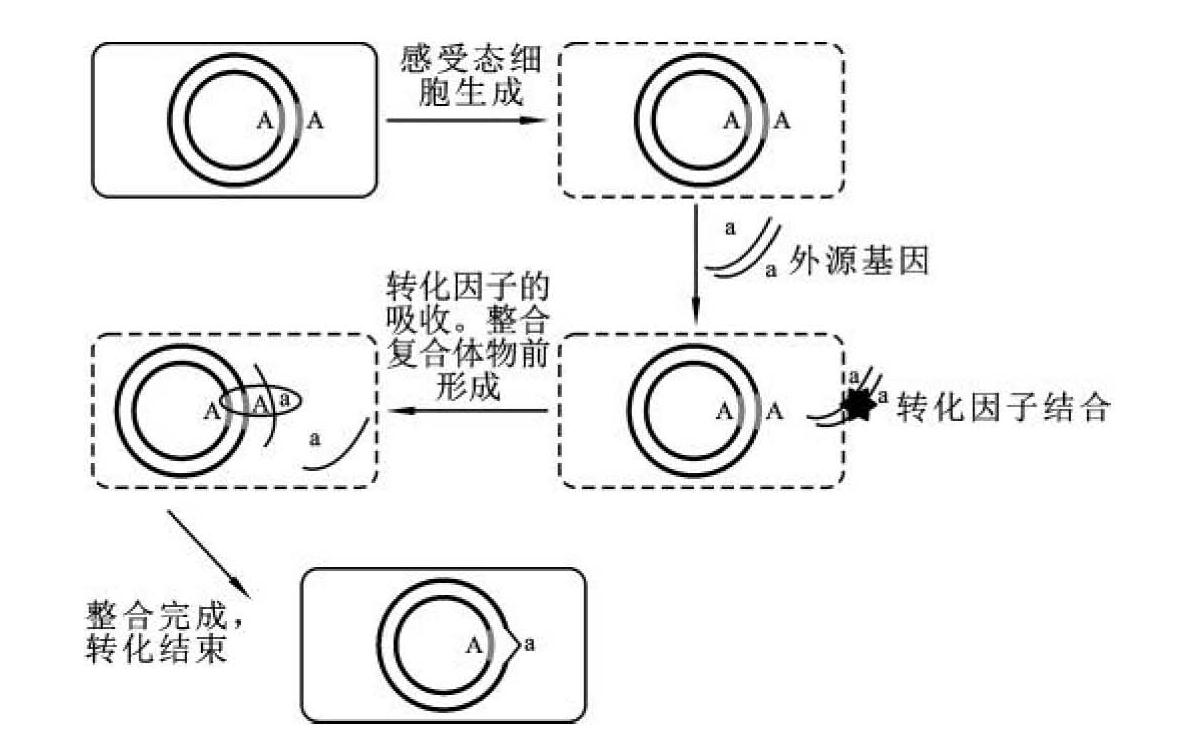
图6-1 转化的基本过程示意图
革兰氏阴性菌的DNA是以完整的双链形式被吸收的,在整合作用发生之前,进入受体细胞内的双链DNA片段与相应的DNA结合蛋白结合,不为核酸酶所降解。DNA整合同样发生在单链水平上,另一条链以及被取代的受体菌单链DNA则被降解。
原核细菌的转化虽是一种较为普遍的遗传变异现象,但是目前仍只是在部分细菌的种属之间发现,如肺炎双球菌、芽孢杆菌、链球菌、假单胞菌以及放线菌等。而在肠杆菌科的一些细菌间很难进行转化,其主要原因是一方面转化因子难以被吸收,另一方面受体细胞内往往存在着降解线状转化因子的核酸酶系统。另外,细菌自然转化是自身进化的一种方式,通常伴随着DNA的整合,因此在DNA重组的转化实验中,很少采取自然转化的方法,而是通过物理方法将重组DNA分子导入受体细胞中,同时也对受体细胞进行遗传处理,使之丧失对外源DNA分子的降解作用,确保较高的转化效率。
2.受体细胞的选择
野生型细菌一般不能用作基因工程的受体细胞,因为它对外源DNA的转化效率较低,并且有可能对其他生物种群存在感染寄生性,因此必须通过诱变手段对野生型细菌进行遗传性状改造,使菌株变成以下类型。
1)限制缺陷型
野生型细菌具有针对外源DNA的限制和修饰系统。如果从大肠杆菌C600株中提取质粒DNA,用于转化大肠杆菌K12株,后者的限制系统便会切开未经自身修饰系统修饰的质粒DNA,使之不能在细胞中有效复制,因此转化效率很低。同样,来自不同生物的外源DNA或重组DNA转化野生型大肠杆菌,也会遇到受体细胞限制系统的降解。为了打破细菌转化的种属特异性,提高任何来源的DNA分子的转化效率,通常选用限制系统缺陷型的受体细胞。大肠杆菌的限制系统主要由hsdR基因编码,因此具有hsdR-遗传表型的大肠杆菌各株均丧失了降解外源DNA的能力,同时大大增加了外源DNA的可转化性。
2)重组缺陷型
野生型细菌在转化过程中接纳的外源DNA分子能与染色体DNA发生体内同源重组反应,这个过程是自发进行的,由rec基因家族的编码产物驱动。大肠杆菌中存在着两条体内同源重组的途径,即RecBCD途径和RecEF途径,前者远比后者重要,但两种途径均需要RecA重组蛋白的参与。RecA是单链蛋白,在同源重组过程中起着不可替代的作用,它能促进DNA分子之间的同源联会和DNA单链交换,RecA-型的突变使大肠杆菌细胞内的遗传重组频率降低至原来的10-6左右。大肠杆菌的recB、recC和recD基因分别编码不同相对分子质量的多肽链,三者构成一个在同源重组中的统一功能RecBCD蛋白(核酸酶V),它具有依赖于ATP的双链DNA外切酶和单链DNA内切酶双重活性,这两种活性也是同源重组所必需的。以外源基因克隆、扩增以及表达为目的的基因工程实验是建立在重组DNA分子自主复制基础上的,因此受体细胞必须选择体内同源重组缺陷型的遗传表型,其相应的基因型为recA-、recB-或recC-,有些大肠杆菌受体细胞则三个基因同时被灭活。
3)转化亲和型
用于基因工程的受体细胞必须对重组DNA分子具有较高的可转化性,这种特性主要表现在细胞壁和细胞膜的结构上。利用遗传诱变技术可以改变受体细胞壁的通透性,从而提高其转化效率。在用噬菌体DNA载体构建的重组DNA分子进行转染时,受体细胞膜上还必须具有噬菌体的特异性吸附受体。
4)遗传互补型
受体细胞必须具有与载体所携带的选择标记互补的遗传性状,才能使转化细胞的筛选成为可能。例如,若载体DNA上含有氨苄青霉素抗性基因(ampicillin resistance gene,Ampr),则所选用的受体细胞应对这种抗生素敏感,当重组分子转入受体细胞后,载体上的标记基因赋予受体细胞抗生素的抗性特征,以区分转化细胞与非转化细胞。更为理想的受体细胞具有与外源基因表达产物活性互补的遗传特征,这样便可直接筛选到外源基因表达的转化细胞。
5)感染寄生缺陷型
相当多的细菌对其他生物尤其是人和牲畜具有感染和寄生效应,重组DNA分子导入这些受体菌中后,极有可能随着受体菌的感染寄生作用进入生物体内,并广泛传播。如果外源基因对人体和牲畜有害,则会导致一场灾难。因此从安全的角度上考虑,受体细胞不能具有感染寄生性。受体细胞选择的另一方面内容是受体细胞种属的确定。对于以改良生物物种为目的的基因工程操作而言,受体细胞的种属没有选择的余地,待改良的生物物种就是受体。但对外源基因的克隆与表达来说,受体细胞种类的选择至关重要,它直接关系到基因工程的成败。
3.转化方法(https://www.daowen.com)
动物细胞和植物细胞的外源DNA导入完整细胞通常采用相应的病毒感染方法。细菌受体细胞的转化方法是基于物理学和生物学原理建立起来的。
1)Ca2+诱导转化
1970年Mandel和Higa发现用CaCl2处理过的大肠杆菌能够吸收噬菌体DNA,此后不久,Cohen等用此法实现了质粒DNA转化大肠杆菌的感受态细胞。将处于对数生长期的细菌置入0℃的CaCl2低渗溶液中,使细胞膨胀,同时Ca2+使细胞膜磷脂层形成液晶结构,使得位于外膜与内膜间隙中的部分核酸酶离开所在区域,这就构成大肠杆菌人工诱导的感受态。此时加入DNA,Ca2+又与DNA结合形成抗脱氧核糖核酸酶(DNase)的羟基-磷酸钙复合物,并黏附在细菌细胞膜的外表面上。经短暂的42℃热激处理后,细菌细胞膜的液晶结构发生剧烈扰动,随之出现许多间隙,致使通透性增加,DNA分子便进入细胞内。此外在上述转化过程中,Mg2+的存在对DNA的稳定性起很大的作用,MgCl2与CaCl2又对大肠杆菌某些菌株感受态细胞的建立具有独特的协同效应。1983年,Hanahan除了用CaCl2和MgCl2处理细胞外,还设计了用二甲基亚砜(dimethyl sulfoxide,DMSO)和二巯基苏糖醇(dithiothreitol,DTT)进一步诱导细胞产生高频感受态的程序,从而大大提高了大肠杆菌的转化效率。目前,Ca2+诱导法已成功地用于大肠杆菌、葡萄球菌以及其他一些革兰氏阴性菌的转化。
2)聚乙二醇介导的细菌原生质体转化
聚乙二醇(PEG),也称为聚环氧乙烷(PEO)或聚氧乙烯(POE),是指环氧乙烷的寡聚物或聚合物。在高渗培养基中生长至对数生长期的细菌,用含有适量溶菌酶的等渗缓冲液处理,剥除其细胞壁,形成原生质体,使之丧失一部分定位在膜上的DNase,有利于双链环状DNA分子的吸收。此时,再加入含有待转化的DNA样品和聚乙二醇的等渗溶液,均匀混合。通过离心除去聚乙二醇,将菌体涂布在特殊的固体培养基上,再生细胞壁,最终得到转化细胞。这种方法不仅适用于芽孢杆菌和链霉菌等革兰氏阳性菌,也对酵母菌、霉菌甚至植物细胞等真核细胞有效。
3)电穿孔驱动的完整细胞转化
电穿孔(electroporation)是一种电场介导的细胞膜可渗透化处理技术。受体细胞在电脉冲的作用下,其细胞壁上形成一些微孔通道,使得DNA分子直接与裸露的细胞膜脂双层结构接触,并引发吸收过程。具体操作程序因转化细胞的种属而异。对于大肠杆菌来说,大约50μL的细菌与DNA样品混合后,置于装有电极的槽内,然后选用大约25μF、2.5 kV和200 Ω的电场强度处理4.6 ms,即可获得理想的转化效率。虽然电穿孔法转化较大的重组质粒(100 kb以上)的转化效率只有小质粒(约3 kb)的约10-3,但这比Ca2+诱导和原生质体转化方法理想,因为这两种方法几乎不能转化100 kb以上的质粒DNA。对于几乎所有的细菌均可找到一套与之匹配的电穿孔操作条件,因此电穿孔转化方法有可能成为细菌转化的标准程序。
4)接合转化
接合(conjugation)是指通过细菌细胞之间的直接接触导致DNA从一个细胞转移至另一个细胞的过程。这个过程是由接合型质粒完成的,它通常具有促进供体细胞与受体细胞有效接触的接合功能以及诱导DNA分子传递的转移功能,两者均由接合型质粒上的有关基因编码(图6-2)。应当特别指出的是,在接合转化过程中使用的重组质粒与接合型质粒必须具有互容性,否则两者难以稳定地存在于供体菌中。
5)噬菌体转染
图6-2 大肠杆菌接合的电子显微镜照片
以DNA为载体的重组DNA分子,由于其相对分子质量较大,通常采取转染的方法将之导入受体细胞内。在转染之前必须对重组DNA分子进行人工体外包装,使之成为具有感染活力的噬菌体颗粒。用于体外包装的蛋白质可以直接从大肠杆菌的溶源株中制备,现已商品化。这些包装蛋白通常分成分开放置且功能互补的两部分,一部分缺少E组分,另一部分缺少D组分。包装时,只有当这两部分的包装蛋白与重组DNA分子三者混合后,包装才能有效进行。任何一种蛋白包装溶液被重组分子污染后均不能包装成有感染活力的噬菌体颗粒,这种设计也是基于安全考虑。整个包装操作过程与转化一样简单:将DNA与外源DNA片段的连接反应液与两种包装蛋白组分混合,在室温下放置1h,加入适量氯仿,离心除去细菌碎片,即得重组噬菌体颗粒的悬浮液。将之稀释合适的倍数,并与处于对数生长期的大肠杆菌受体细胞混合涂布,过夜培养,即可用于筛选与鉴定。
4.影响转化率的因素
转化率的高低与一般重组克隆实验关系不大,但在构建基因文库时,保持较高的转化率至关重要。影响转化率的因素很多,包括以下三个方面。
(1)载体DNA及重组DNA:载体本身的性质决定了转化率的高低,不同的载体DNA转化同一受体细胞,其转化率明显不同。载体分子的空间构象对转化率也有明显影响,超螺旋结构的载体质粒往往具有较高的转化率,经体外酶切连接操作后的载体DNA或重组DNA由于空间构象难以恢复,其转化率一般要比具有超螺旋结构的质粒低两个数量级。对于以质粒为载体的重组分子而言,相对分子质量大的转化率低,到目前为止,30 kb以上的重组质粒很难进行转化。
(2)受体细胞:受体细胞除了具备限制重组缺陷性状外,还应与所转化的载体DNA性质相匹配,如pBR322转化大肠杆菌JM83株,其转化率不高于103/g(DNA),但若转化ED8767株,则可获得106/g(DNA)的转化率。
(3)转化操作:受体细胞的预处理或感受态细胞的制备对转化率影响最大。对于Ca2+诱导的完整细胞转化而言,菌龄、CaCl2处理时间、感受态细胞的保存期以及热激时间均是很重要的因素。其中感受态细胞通常在12~24 h内转化率最高,之后转化率急剧下降。对于原生质体转化而言,再生率的高低直接影响转化率,而原生质体的再生率又受诸多因素的制约。在一次转化实验中,DNA分子数与受体细胞数的比值对转化率也有影响,通常50~100 ng的DNA对应于108个受体细胞或原生质体,在此条件下,加大DNA量并不能线性提高转化率,甚至反而使转化率下降。不同的转化方法导致不同的转化率,这是不言而喻的。其中电穿孔法的转化率与质粒大小密切相关,但明显优于Ca2+诱导的转化,接合转化虽然转化率较低,但对于那些不能用其他方法转化的受体细胞来说不失为一种选择,如光合细菌大多数种属的菌株均采用接合转化方式将重组DNA分子导入细胞内。
5.转化细胞的扩增
转化细胞的扩增是指受体细胞经转化后立即进行短时间的培养,如Ca2+诱导转化后的受体细胞在37℃培养1h、原生质体转化后的细胞壁再生过程以及重组DNA分子体外包装后与受体细胞的混合培养等。转化细胞的扩增主要包括:①转化细胞的增殖,使得有足够数量的转化细胞用于筛选环节;②载体DNA上携带的标记基因拷贝数扩增及表达,这是进行筛选操作的前提;③克隆的外源基因的表达,如果重组DNA分子的筛选与鉴定依赖于外源基因表达产物的检测,则外源基因必须在转化细胞扩增期间表达。总之,转化细胞扩增的目的只有一个,即为后续的筛选鉴定创造条件。