3.3.3 单链DNA噬菌体载体
在以大肠杆菌为宿主细胞的噬菌体家族中,λ和T系列噬菌体为双链结构相对分子质量较大的噬菌体,同时还存在着相对分子质量较小的单链环形结构的噬菌体。M13噬菌体就是一种典型的单链DNA小相对分子质量的噬菌体,其特异性感染含有F性须结构的大肠杆菌,此外fl和fd噬菌体也是和M13噬菌体同源性很高的单链DNA噬菌体。
1.M13噬菌体的结构
并非所有噬菌体都含有多面体的头部外壳结构,例如丝状噬菌体就是一种长度可变、伸展、柔软可弯曲的管道状结构噬菌体,这种长度可变的结构可以包装不同大小的DNA分子。M13噬菌体是一种典型的丝状噬菌体。M13噬菌体呈长而柔软的圆筒状(图3-19(a)),长度达到圆筒结构直径的150倍。这种筒状结构由基因8编码的P8蛋白以超螺旋形式排列组装而成,每个P8蛋白与M13噬菌体筒状结构以一个较小的夹角向外伸展(图3-19(b)),由5个P3蛋白组成复合体构成M13噬菌体的一端,P3蛋白是一种较P8蛋白更大而柔软的蛋白质结构,在感染宿主细胞时P3蛋白可以和宿主细胞表面特定受体结合。筒状结构相反的另一端由P7和P9蛋白组成,这一结构参与M13子代噬菌体从宿主细胞释放的过程。在筒状外壳内部包裹着M13噬菌体单链环状DNA(图3-20)。成熟的M13噬菌体内只含有由DNA(一)转录出来的DNA(+)(也称为感染性单链),DNA含有6407 bp,以重叠基因形式编码11种蛋白质(图3-20)。

图3-19 M13噬菌体外壳结构
(引自Slonczewski J.L.与Foster J.W.,2011)
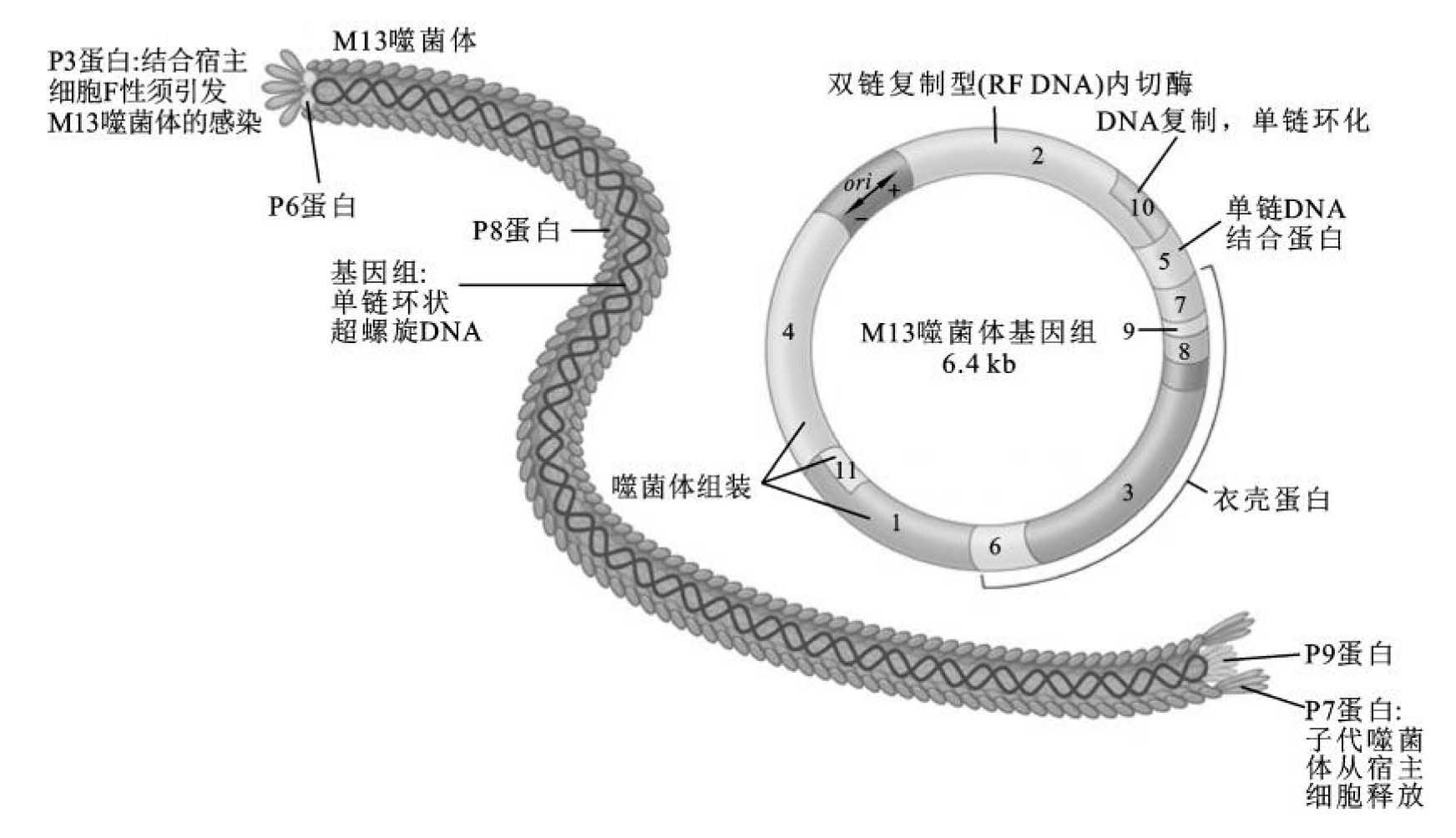
图3-20 M13噬菌体结构及基因组
(引自Slonczewski J.L.与Foster J.W.,2011)
2.M13噬菌体的增殖过程
M13噬菌体只能感染有F性须的雄性大肠杆菌,P3蛋白对宿主细胞F性须的识别与结合在噬菌体的感染吸收过程发挥重要作用(图3-21)。P3蛋白和发挥第一受体作用的F性须接触导致F性须收缩,这一过程引发P3蛋白和宿主细胞第二受体TolA蛋白结合。TolA复合体协助M13噬菌体和宿主细胞的紧密结合。伴随M13噬菌体丝状结构和宿主细胞的紧密结合,M13噬菌体DNA跨过细胞膜被导入宿主细胞。目前,这一过程的具体机制仍不明确。和λ和T系列噬菌体不同,M13噬菌体的外壳不会留在宿主细胞外,构成筒状结构的蛋白亚基分解后进入宿主细胞,参与子代M13噬菌体组装。
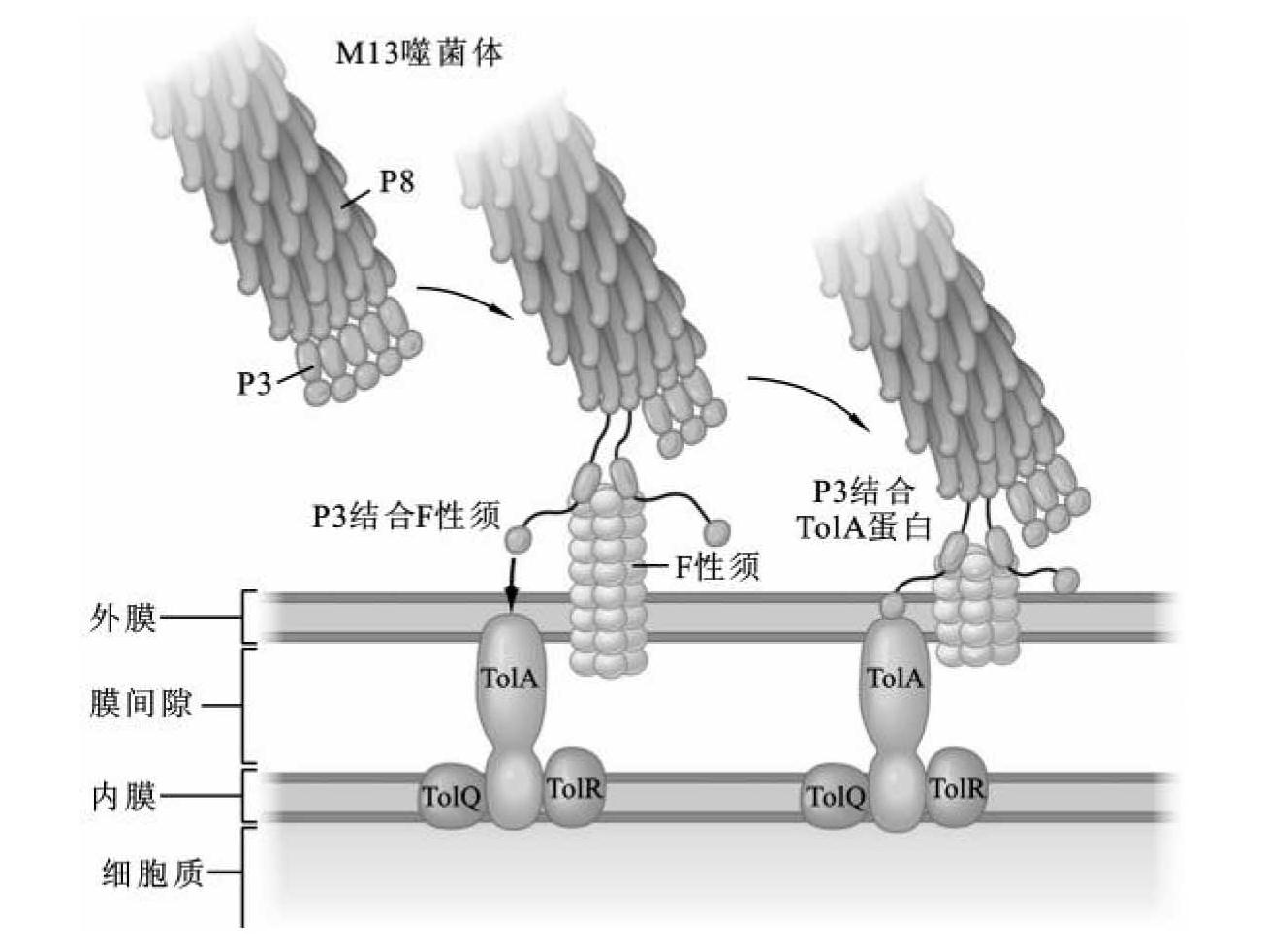
图3-21 M13噬菌体与宿主细胞F性须的结合
(引自Slonczewski J.L.与Foster J.W.,2011)
M13噬菌体DNA(+)进入宿主细胞质后,伴随宿主细胞的增殖M13 DNA利用细胞内的酶进行复制。M13 DNA(+)先合成出DNA(一)形成环形双链结构(图3-22(a)),此结构称为复制型DNA(replication form DNA,简称RF DNA)。RF DNA首先进行几轮“0”复制,使RF DNA的数量达到大约200个拷贝,随后M13噬菌体基因Ⅱ产物结合在RF DNA(+)特定位点并切割出一个缺口。此后,利用宿主细胞内的DNA聚合酶Ⅰ以环形M13 DNA(-)为模版持续合成M13 DNA(+)。M13 DNA基因5编码P5蛋白与新合成的DNA(+)结合,发挥保护DNA的作用,而且认为P5蛋白还参与不对称的DNA复制过程,即只复制M13 DNA(+)而不复制DNA(-)。P5蛋白作为单链DNA特异性结合蛋白,和DNA(+)结合,从而阻断以DNA(+)为模版合成DNA(-)。
结合有P5蛋白的环化的M13 DNA(+)被运送到宿主细胞膜位置包装为子代噬菌体。M13基因组提前合成出子代噬菌体组装所需要的结构蛋白和调控蛋白,如衣壳结构P8蛋白和组成尾部结构的P3、P7和P9蛋白,以及调控子代噬菌体组装的P4蛋白等。P4蛋白单体在细胞膜位置组装为贯通细胞内膜和外膜的圆筒状包装装置,子代M13噬菌体在孔道结构内完整组装。为了防止宿主细胞被裂解,在子代M13噬菌体组装开始之前,P4蛋白组成的包装装置孔道会被P1和P11蛋白封闭。组装过程如图3-22(b)所示,首先,带有正电荷的P4蛋白N末端结构会吸附带有负电荷的M13 DNA(+)进入包装装置,在包装装置入口处,P5蛋白被卸下,带有正电荷的P8蛋白被逐个组装到M13 DNA分子上,子代噬菌体释放的最后一步是组装P3蛋白。当子代噬菌体释放后,P4组装装置的孔道会再次被P1和P11蛋白封闭,防止宿主细胞被裂解。在此包装过程中,M13噬菌体颗粒的大小和被包装的M13 DNA大小有关,因此M13噬菌体包装DNA的相对分子质量要求相对宽松,最大的包装容量可以达到野生型M13噬菌体DNA相对分子质量的7倍。每个宿主细胞在M13噬菌体繁殖周期可以释放约1000个子代噬菌体颗粒。M13菌体的增殖过程不裂解宿主细胞,但会导致宿主细胞增殖速度减慢。在M13 DNA中编码的11个基因中有2个基因是防止降解宿主细胞的,这种进化策略使其可以获得更多后代。这种非溶菌性增殖方式在基因工程应用上有很大的优势,可以在培养基中聚集大量M13噬菌体颗粒,便于子代噬菌体分离纯化。
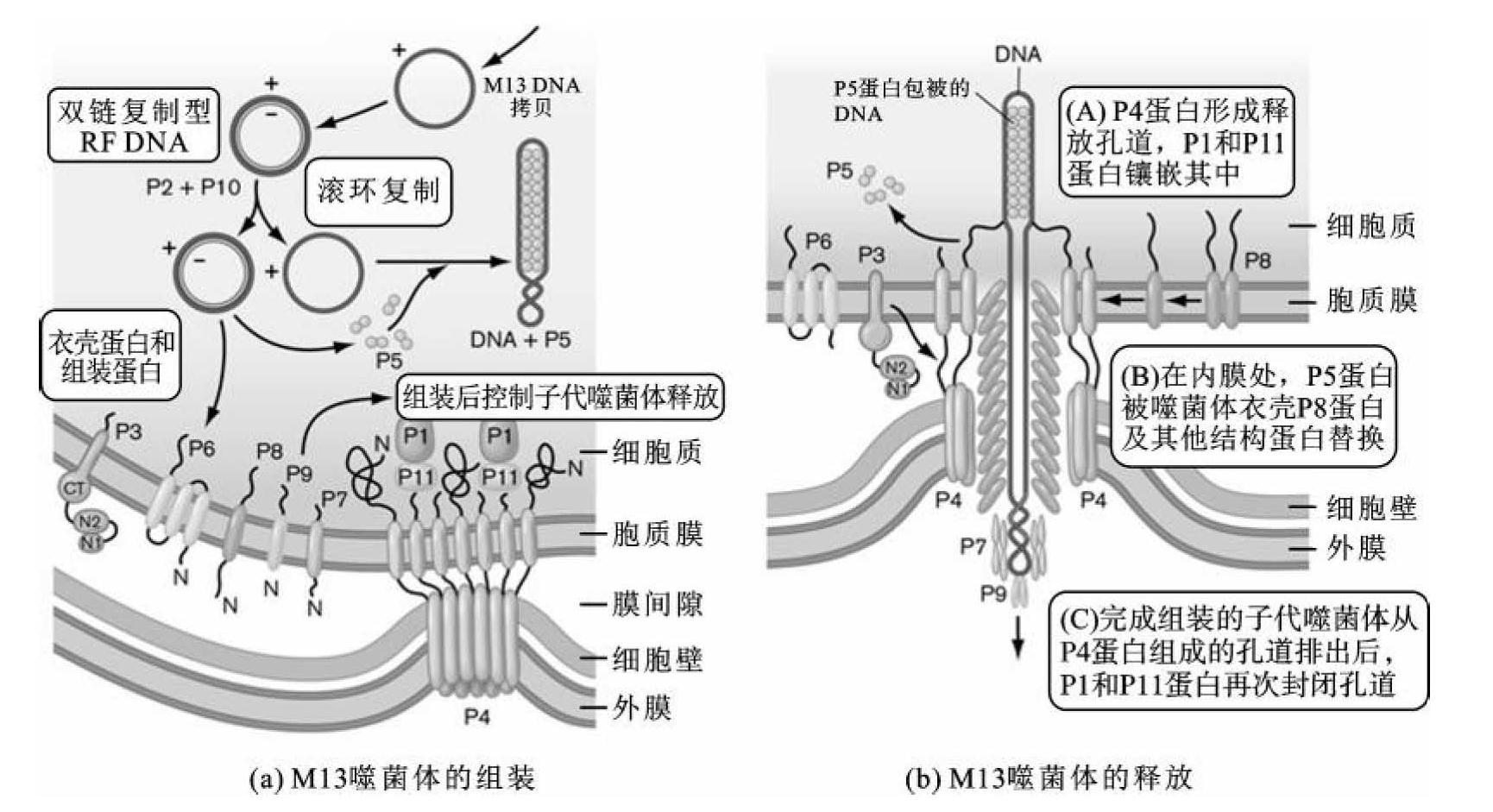
图3-22 子代M13噬菌体的组装与释放
(引自Slonczewski J.L.与Foster J.W.,2011)
3.野生型M13噬菌体的改造
在对野生型M13噬菌体进行改造的过程中,首先确定了其DNA中可用的克隆区域。研究发现,几乎所有的M13噬菌体上的基因都非常重要,不存在类似λDNA的非必需区段,只在基因Ⅱ和Ⅳ之间存在507 bp的区间可以进行外源DNA插入。对M13噬菌体DNA改造过程如下:
(1)引入遗传标记基因,将β-半乳糖苷酶α片段基因(lac Z')及其配套的操纵子元件引入M13 DNA中,便于利用蓝白斑筛选M13 DNA重组体,引入抗生素抗性筛选标记,便于转化子筛选;
(2)利用定点诱变技术去除在M13 DNA中多次出现的酶切位点,并在基因Ⅱ和Ⅳ间插入人工合成的多克隆位点(MCS)接头,MCS区域位于lac Z'中,便于外源DNA的克隆和重组体的蓝白斑筛选;(https://www.daowen.com)
(3)在MCS两侧加入测序引物互补序列,便于将提取的M13 DNA直接用于测序。
目前常用的M13-DNA克隆载体为M13mp系列载体,其中M13mpl0和M13mpll的结构如图3-23所示,这两种M13噬菌体载体带有一段限制性位点相同但方向相反的MCS区域。因此任何一种可以插入MCS区域的外源DNA片段,都能够按两种彼此相反的取向进行克隆。这一特点对于DNA序列分析非常有用,可以同时从两个方向对同一段外源DNA测序。
4.单链噬菌体DNA载体的优势
丝状噬菌体结构简单,内部DNA呈单链环状结构,噬菌体DNA复制过程既包含类似于质粒DNA的双链结构阶段,又包含子代噬菌体DNA的单链环状结构阶段,这些特殊的特征使丝状噬菌体具备了成为基因工程载体和研究工具的优势:
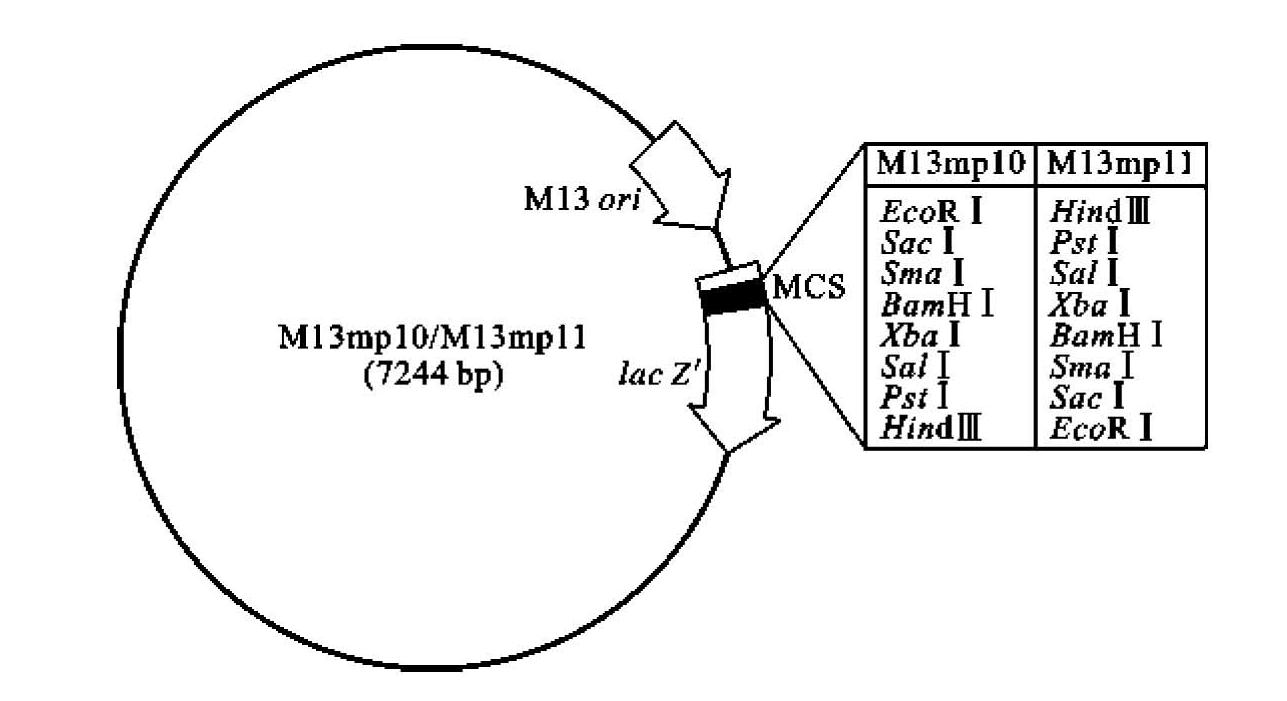
图3-23 M13克隆载体M13mpl0和M13mpl1结构图
(1)非溶菌性增殖方式可以在培养基内收集大量的子代噬菌体颗粒,便于以类似于λDNA的方式进行DNA纯化;
(2)子代噬菌体DNA只含正链DNA(+),其中包含克隆DNA片段的两条互补链中的一条,因此可直接用于DNA测序和定点诱变,或用于制备单链DNA探针以选择和分离互补的RNA;
(3)单链噬菌体DNA的RF型是双链DNA,便于以类似质粒的方法进行分离纯化,且限制性核酸内切酶可以对其识别和切割,便于外源DNA与双链RF DNA的酶切和连接;
(4)当克隆的外源DNA相对分子质量不是很大时,便于以质粒转化的方式进入宿主细胞,不需要进行体外包装。
M13噬菌体载体在使用过程中也存在一些缺点:第一,外源DNA插入会降低M13噬菌体载体稳定性,插入片段越大,则载体稳定性越低;第二,虽然理论上外源DNA片段可以按两个方向插入M13噬菌体载体,但研究发现,它经常以一种主要的方向插入酶切位点;第三,M13噬菌体载体承载能力差,虽然理论上M13噬菌体载体无包装的限制,但实际情况下,通常插入的最大片段仅1500 kb。由单链噬菌体载体和质粒载体融合构建的人工载体对这些缺点有明显的改善。