3.7.3 酵母人工染色体载体
Murray和Szosta在1983年将酵母的着丝粒、ARS序列及四膜虫核糖体RNA基因rDNA(Tr)末端序列插入大肠杆菌质粒pBR322中,并转化酵母菌,构建成第一个酵母人工染色体,进一步改造使YAC能够在后代中稳定传递。由于当时人类基因组计划(Human Genome Project)需要每条染色体的高分辨率物理图谱,而且急需一种能够将染色体变成小片段进行直接测序的大片段DNA载体,因此,YAC被应用于人类基因组计划。通常YAC载体能够容纳200~500 kb DNA片段,最高可以达到1 Mb染色体片段。YAC重组体导入宿主细胞后通常采用营养缺陷型标记基因进行重组体筛选,如色氨酸、亮氨酸和组氨酸合成缺陷型基因TRP1、LEU2和HIS3,尿嘧啶合成缺陷型基因URA3,以及赭石突变抑制基因SUP4。酵母从头合成腺嘌呤核苷酸和次黄嘌呤一磷酸时需要磷酸核糖胺咪唑羧化酶,编码该酶的基因ADE2有一种突变形式,称为ADE2-1,其内部第64位密码子由GAA变成赭石型终止密码子TAA,所以称为赭石型突变,它导致翻译终止,不能产生有活性的磷酸核糖胺咪唑羧化酶,酵母细胞内合成嘌呤的中间产物磷酸核糖胺咪唑聚合成红色物质,使ADE2-1酵母菌落呈红色。SUP4基因是一种反密码子发生突变的酪氨酸tRNA基因,能够阅读宿主酵母的ADE2-1内部的TAA,使翻译继续,产生有功能的磷酸核糖胺咪唑羧化酶,抑制了ADE2-1的突变效应,使ADE2-1酵母菌落呈白色,因此被称为ADE2-1的赭石型抑制子。将YAC载体中SUP4基因区段作为外源DNA插入区域。外源DNA的插入导致SUP4失活,失去抑制赭石型突变的能力,从而产生红色菌落。
多种常用的酵母人工染色体载体都是在pYAC3和pYAC4两种YAC载体基础上改进而来,因此下面以pYAC3为例简要介绍酵母YAC载体的结构与功能。pYAC3(图3-29)载体呈双链环状,承载了β-内酰胺酶基因(β-lactamase,bal),可以进行β-内酰胺类抗生素筛选,如氨苄青霉素(Amp)筛选,并带有细菌pMB1质粒的复制起始位点,可以在细菌细胞中自主复制。同时,其内部还带有酵母复制起始位点ARS1序列和着丝粒CEN4序列,保证其在酵母细胞内的复制和分离。利用TRP1、URA3和HIS3营养筛选标记基因和赭石突变抑制基因SUP4对重组体在酵母细胞内进行营养筛选和菌落颜色筛选。从四膜虫基因组中克隆的两个端粒样DNA序列TEL使pYAC3在酵母细胞中可以形成端粒结构,保证了YAC在细胞内的稳定性。
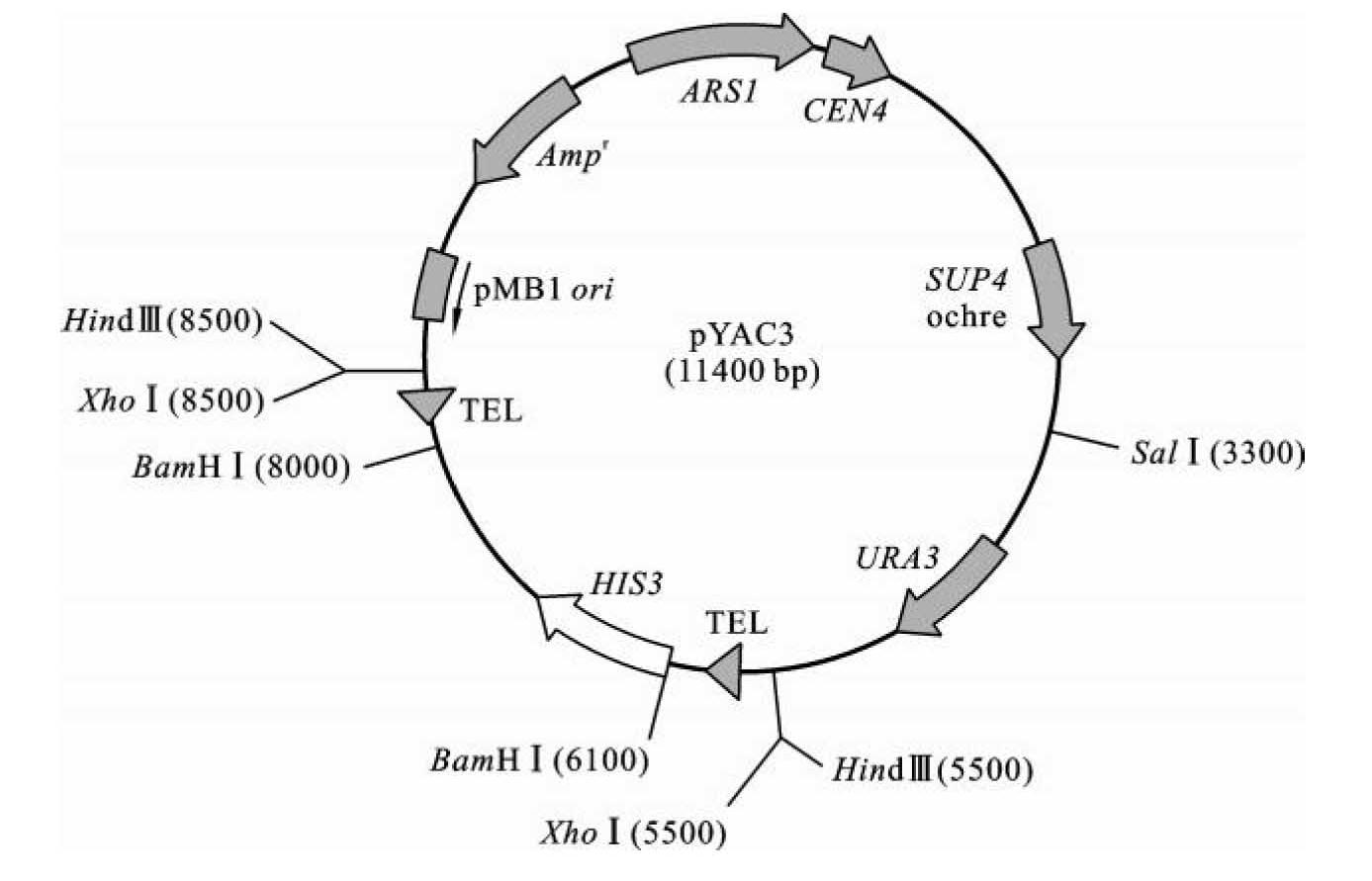
图3-29 酵母人工染色体载体pYAC3结构
外源DNA片段与pYAC3的重组过程如下(图3-30):
(1)利用HIS 3筛选标记两端的Bam HⅠ酶切位点,将pYAC3载体线形化,此过程导致HIS 3被切除,因此可以利用HIS 3营养标记基因对切割不完全的载体进行逆向筛选;
(2)利用SnaB Ⅰ限制性核酸内切酶将SUP4内部Sna B Ⅰ酶切位点断开,形成外源DNA片段插入位点,SnaB Ⅰ限制性核酸切割产物为平末端的两条线状YAC载体的左臂和右臂;
(3)利用连接酶将外源DNA片段同磷酸酶处理的YAC左臂和右臂连接,转入宿主细胞进行三轮筛选。(https://www.daowen.com)
pYAC3重组体转入带有URA3突变、TRP1突变和赭石反应突变基因ADE2-1的宿主酵母菌(如AB1380),筛选原理要点如下:
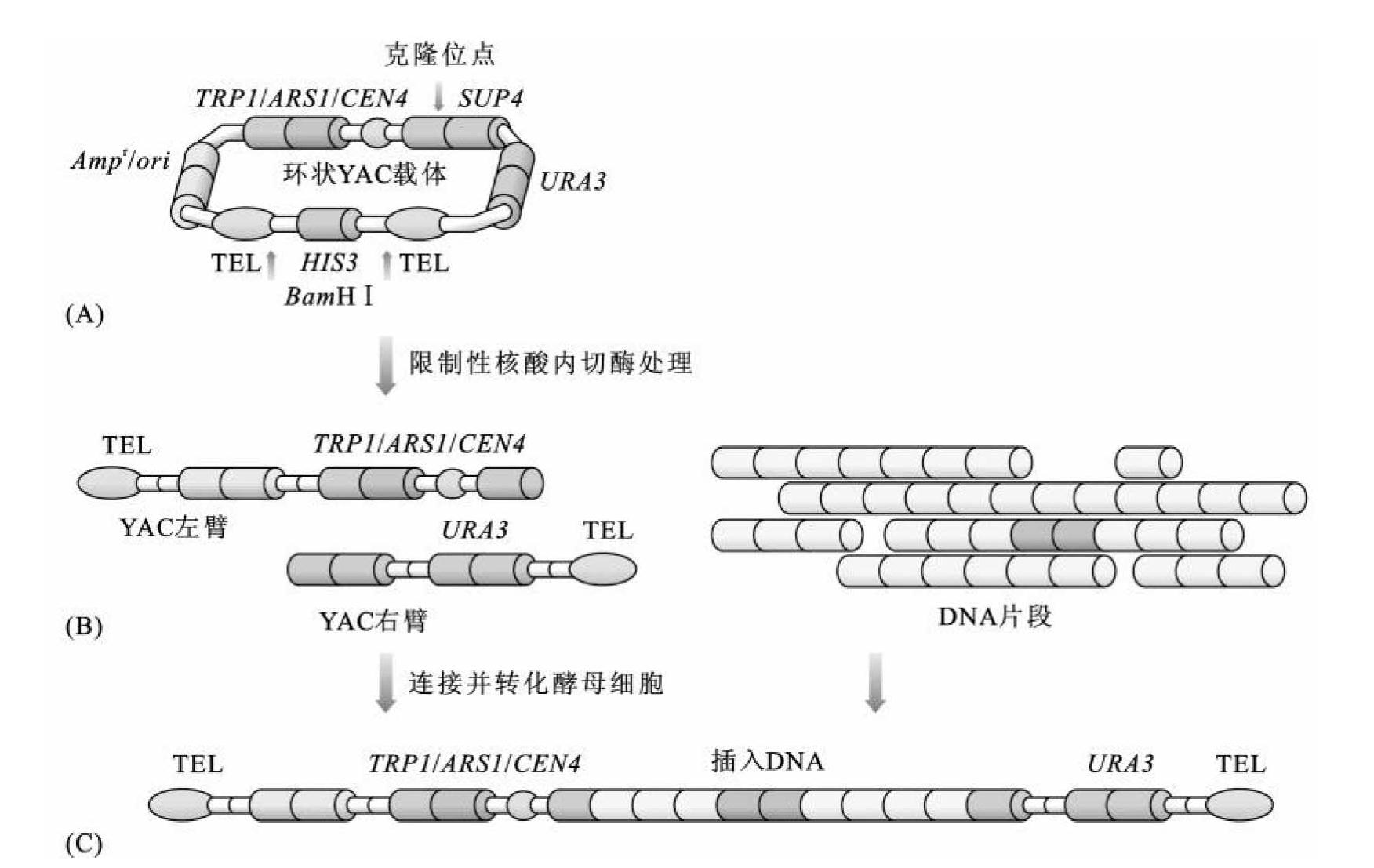
图3-30 外源DNA片段和pYAC3重组过程
①利用宿主酵母URA3突变和载体右臂UAR3产物互补的原理,进行营养缺陷筛选,筛选含有YAC右臂的重组体;
②利用宿主酵母TRP1突变和载体左臂TRP1产物互补的原理,进行营养缺陷筛选,筛选含有YAC左臂的重组体;
③将外源DNA插入载体,导致SUP4失活,宿主生成红色菌落,利用颜色反应筛选插入外源DNA片段的pYAC3重组体。
目前,在人类、小鼠、果蝇、拟南芥和水稻等高等生物中均构建了高质量YAC文库。然而,YAC具有一些缺陷。例如,存在高比例嵌合体,即一个YAC克隆含有两个本来不相连的独立片段;部分克隆不稳定,在传代培养中可能发生缺失或重排;因为YAC与酵母染色体具有相似的结构,因此难以与酵母染色体分离;YAC在细胞中以线状存在,操作时容易发生染色体机械切割。