8.2.4 在酵母中高效表达外源基因的策略
外源基因在宿主细胞内的有效表达是基因工程的核心问题之一,利用宿主菌合成某种真核生物的蛋白质以满足商品生产的广泛需求,仅仅停留在实验室检测水平上的表达量是远远不够的。因此,使外源基因高效表达是急需解决的问题。使用酵母表达真核基因获取蛋白质产物是基因工程中较为成熟的生产手段,人们已在筛选强启动子、调控因子以及提高表达载体稳定性和拷贝数等方面做了大量的工作,其表达外源基因主要有以下几种策略。
1.构建合适的表达菌株
要使外源蛋白在酵母中得到高效表达,首先是要构建合适的表达菌株。一般而言,蛋白质的合成产量与基因的拷贝数成正比,因此增加外源基因的拷贝数是常见的一种用于提高外源基因表达效率的方法。要获得高效稳定的外源蛋白表达菌株,首先要将毕赤酵母表达载体连同外源基因通过同源重组途径整合到宿主染色体基因组上。根据整合时线形化位点的不同,分为两种整合方式,即单插入和置换,产生两种不同的表型。单插入通常导致插入不同拷贝数的基因组,而置换一般是将单拷贝外源基因整合入基因组。整合型表达载体与自主复制的质粒型表达载体有不同的特点,其拷贝数可以有很大的变化。含有多个拷贝的外源基因时,表达菌株合成蛋白质的量也会显著增大。然而多拷贝的外源基因也具有一定的缺点:多拷贝的发生也会干扰重组子的筛选;目的基因拷贝数较多,会给宿主菌造成内质网压力而降低表达。因此适当的外源基因拷贝数有利于外源蛋白的表达,当前主要是通过人工构建的方式得到合适的表达载体,然后整合到宿主菌获得多拷贝表达菌株。主要有两种方法可获得多拷贝表达菌株:一种是在大量的重组转化子中利用免疫杂交或菌落斑点印迹杂交方法筛选产量高的表达菌株;另一种是在转化前先将多个表达盒拷贝插入单个载体中,而后通过同源重组整合到受体染色体上。以上两种方法要在发酵罐培养的产物选择压力下,才能获得稳定的多拷贝表达菌株。此外,利用内源性质粒也可获得多拷贝质粒载体,但其稳定性无法保持。也可利用酵母rDNA的多拷贝基因来介导载体的多拷贝整合,使外源基因得到高水平表达。
利用毕赤酵母作为宿主细胞表达乙肝表面抗原(HBsAg)时(图8-8),比酿酒酵母效果更好。将HBsAg的编码序列和选择标记his4(组氨酸脱氢酶基因)插入AOX1的启动子和终止子之间获得pBSAG151重组质粒。重组质粒用Bgl Ⅱ线形化后,AOX1的启动子和终止子分别位于两端。用HIS-受体细胞选择HIS+转化子,HBsAg的编码序列通过同源重组整合到宿主染色体上。未受影响的AOX2使转化子可在甲醇培养基中生长。重组菌产生的S蛋白可达细胞可溶性蛋白的2%~3%,比含有多拷贝表达单元的重组酿酒酵母高近1倍。
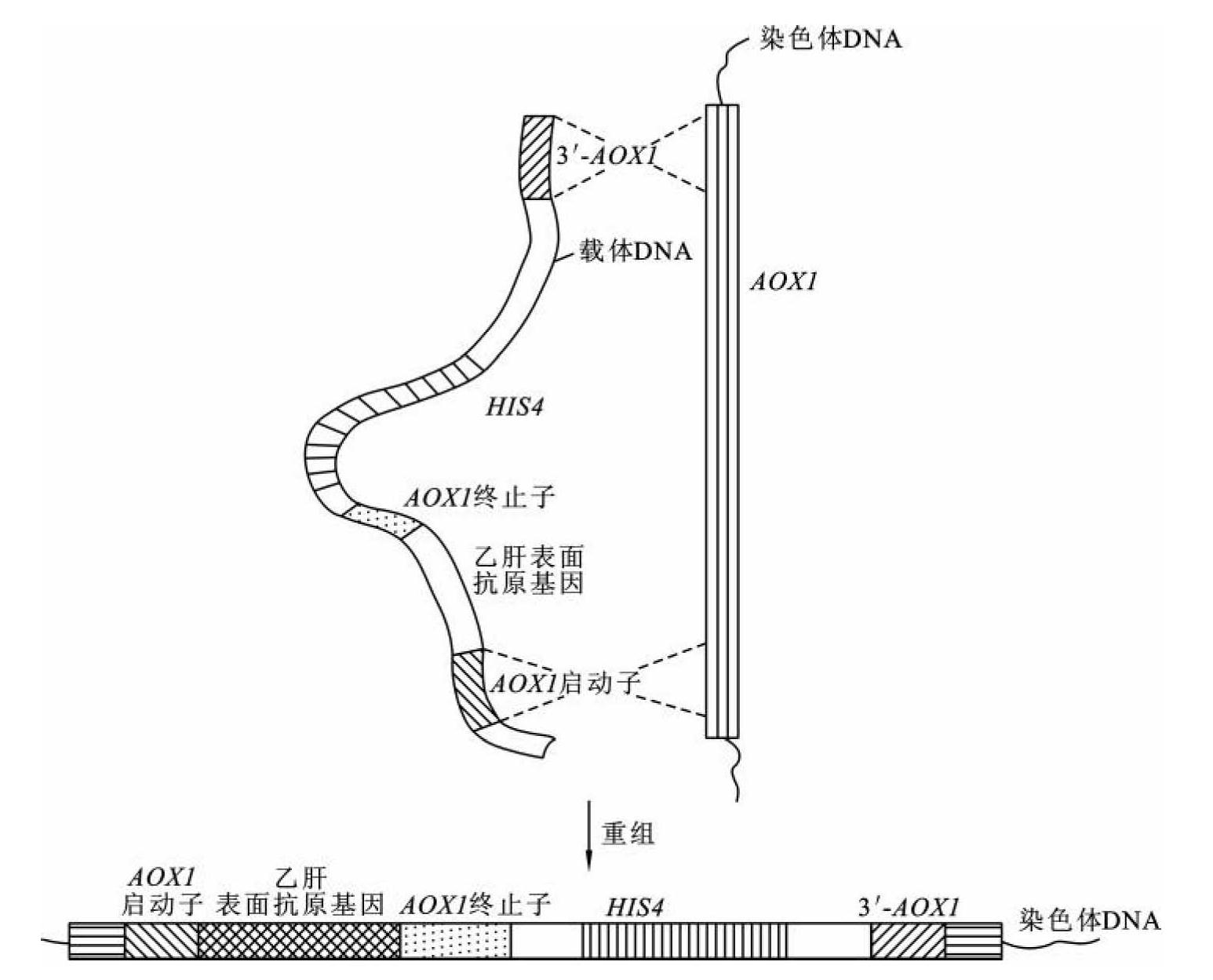
图8-8 毕赤酵母中乙肝表面抗原的表达
(引自瞿礼嘉等,1998)
2.外源基因的特征
不是任何基因都适合在酵母中进行表达,外源基因的自身特征也是影响基因表达水平的重要因素。如将蜘蛛的丝蛋白基因整合到表达载体中在酵母中进行表达,当拷贝数逐渐增加时,其生产效率却有所降低,这与丝蛋白基因自身的特征密切相关。此外,在许多富含AT的基因中容易出现提前终止转录的现象,使外源基因的表达效率受到限制,不能有效地转录并翻译形成蛋白质产物。相反,基因中GC含量过高,特别是在mRNA 5'翻译区,则容易导致翻译能障过高,翻译过程不能顺利进行,进而导致表达量的减少。因此,可以通过调整外源基因的AT和GC含量来使外源基因顺利表达。而mRNA 5'非翻译区的核苷酸序列长度不合适也可能影响到基因的表达,成为外源基因高效表达的限制因素。转录提前终止被认为是一种具有种属特异性的现象,如HIV ENV蛋白在毕赤酵母中不能表达,但在啤酒酵母中表达良好。因此,可以通过对高AT含量区的核苷酸组成进行调整来避免转录提前终止的发生。Sreekrishna等通过研究发现,将人血白蛋白(human serum albumin,HAS)的mRNA 5'非翻译区的核苷酸组成进行调整,使其与醇氧化酶的5'非翻译区的核苷酸序列相同,则HAS的表达量可以提高50倍以上。但遗憾的是,限于目前对毕赤酵母的了解程度,仍然无法预见某种外源蛋白能否在其中表达或获得高产量。(https://www.daowen.com)
3.提高表达产物质量
酵母具有多种蛋白质修饰能力,如乙酰化修饰、甲基化修饰等,另外酵母还有较强的亚基装配能力,如哺乳动物钠钾ATP酶的α、β亚基以及人胚胎血红蛋白的α、β亚基在酵母中能进行表达和装配。酵母可以对很多外源蛋白进行分泌表达,而分泌表达不仅能提高表达水平,简化产物纯化步骤,还可使产物通过分泌途径完成加工和修饰。
4.其他因素
还可以考虑通过优化其他因素来提高表达效率,包括:提高翻译效率,如优化翻译起始区前后的mRNA二级结构,在外源基因中尽量选用酵母偏爱的密码子;避免表达产物在细胞内降解,可以使用蛋白酶缺失的菌株;优化工程菌发酵工艺,如进行高密度发酵、控制发酵参数和发酵时间。