8.2.2 酵母基因表达载体的种类
酵母基因表达载体可以分为三种,即酵母质粒载体、酵母整合载体、酵母人工染色体。
1.酵母质粒载体
酵母质粒载体广泛用于表达胞内或胞外的重组蛋白,但游离的质粒表达系统不够稳定。当酵母质粒载体整合到宿主染色体上以后稳定性增加,由于整合后的克隆基因拷贝数仅为1,因而外源基因表达效率低。基因串联可以增加外源基因拷贝数和表达产量,但质粒稳定性会受影响。
酵母质粒载体分为酵母附加型质粒(yeast episomal plasmid,YEp,图8-1)、酵母复制质粒(yeast replicating plasmid,YRp,图8-2)和酵母着丝粒质粒(yeast centromere plasmid,YCp,图8-3)。
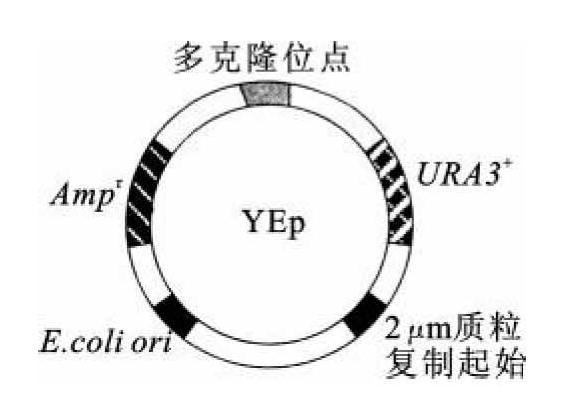
图8-1 YEp质粒图谱
(引自张惠展,2005)
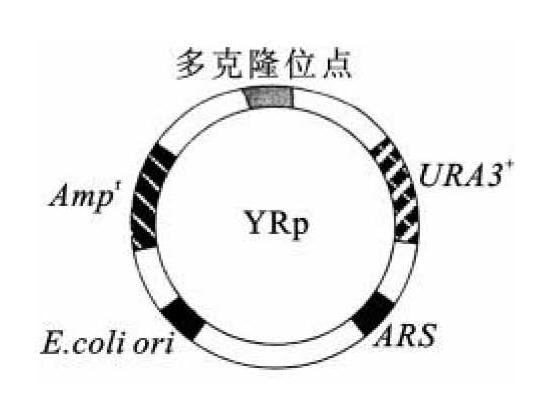
图8-2 YRp质粒图谱
(引自张惠展,2005)
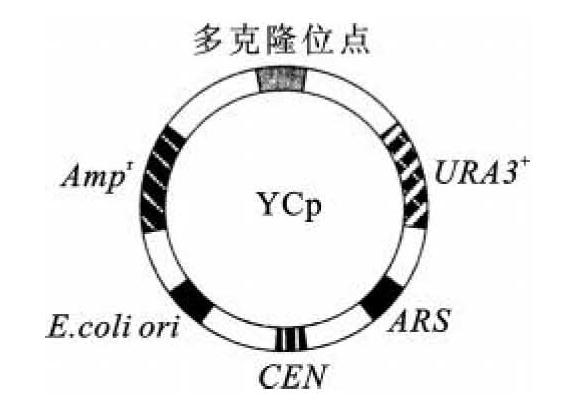
图8-3 YCp质粒图谱
(引自张惠展,2005)
酵母附加型质粒是酵母的野生型质粒,大小为6.3 kb,长约2μm,称为2μm质粒。在一个单倍体细胞中其拷贝数可达50~100,占到酵母基因组的2%~4%。野生型质粒的一个重要特点是,该质粒被一段长度为599 bp的反向重复序列分为两部分,每部分都含有一个启动子和该启动子控制下的两个基因。其中的一个基因称为FLP,它编码一个位点特异性的重组酶(recombinase),这个酶可以催化反向重复序列之间的遗传重组。这样2μm质粒就有两种存在形式(图8-4)。YEp就是在2μm质粒的基础上,加入酵母核DNA序列,以及大肠杆菌质粒pMB9的部分序列组成的。这样,YEp就既可以在酵母细胞中复制,也可以在大肠杆菌中复制。
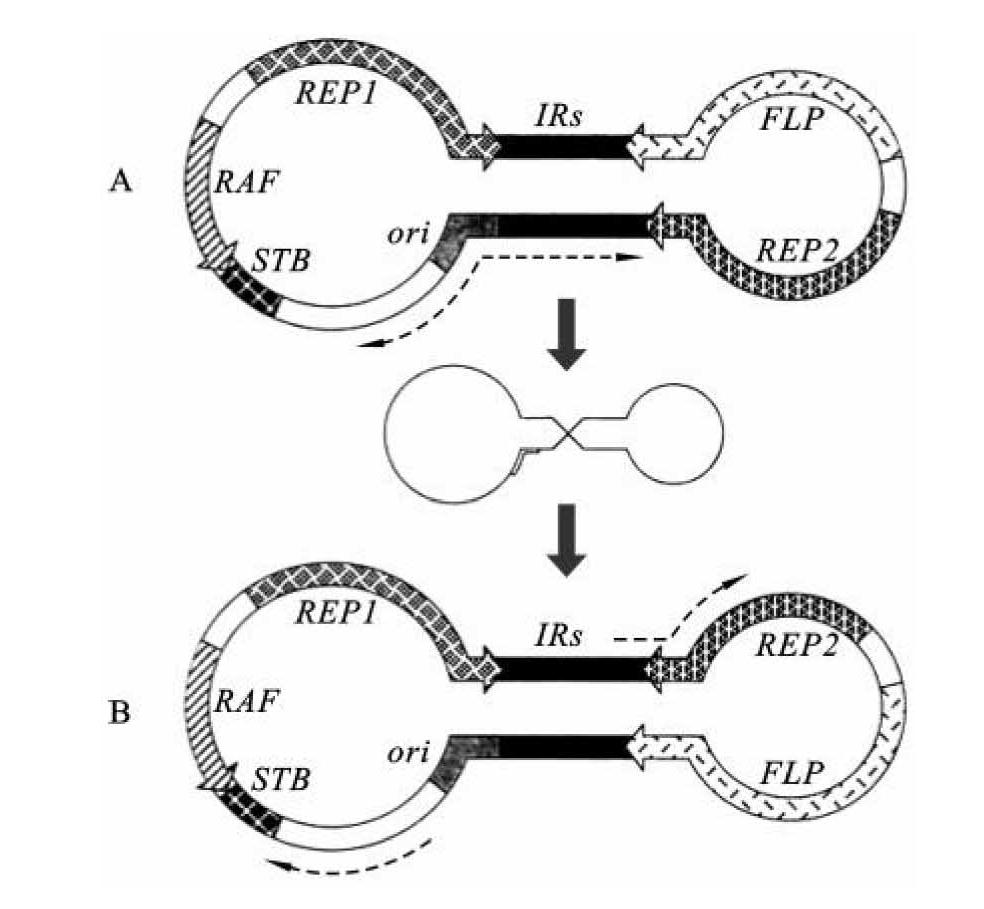
图8-4 2μm双链环状质粒的两种不同形态
(引自张惠展,2005)
酵母复制质粒是大肠杆菌质粒pBR322的衍生质粒。在pBR322中加入一段来源于酵母染色体的自主复制序列后,质粒就可以在酵母中进行复制。酵母复制质粒一般不会与酵母染色体发生重组,其转化效率很高,但是不能稳定地传代,这是此载体的一个严重缺点。
酵母着丝粒质粒克服了酵母复制质粒不能稳定传代的缺点。在酵母的3号染色体着丝粒周围的leu2和cdc10之间的一段1.6 kb序列可以稳定遗传,并含有2个自主复制序列。在4号和9号染色体上也存在功能相同的序列。带有此着丝粒序列的这类质粒在酵母中的行为类似于微型染色体,在宿主细胞中的拷贝数低,可以稳定遗传,均匀分配给子代细胞。
2.酵母整合载体
酵母整合载体(yeast integration plasmid,Yip)是指不含酵母自主复制序列,因而不能在酵母中独立复制的一种载体。这种载体必须在整合到酵母染色体中之后才能使其中所包含的基因组得到稳定表达。它带有整合介导区,可通过同源重组整合到酵母基因组中与其同步复制。
在啤酒酵母的基因组中的可动遗传因子——Ty因子,与逆转录病毒在结构和功能上有很多相似之处,包括两个长的ORF和两个长末端重复序列(LTR),这两个长末端重复序列称为δ因子。基因组中含有30~40个拷贝的Ty,由于Ty可以整合到宿主细胞的染色体中,适合于构建整合载体。经过研究发现,用磷酸甘油酸激酶启动子代替δ因子中原有的启动子序列,可以使Ty的拷贝数大大提高,从而在细胞质中形成大量的类似病毒颗粒(virus-like particle,VLP)的物质。
毕赤酵母没有稳定的附加体质粒,其表达载体一般是整合型载体。毕赤酵母表达系统最早在20世纪90年代由Phillip Petrocleum公司的研究人员开发,该表达系统利用AOX1基因的启动子,使整合到AOX1基因启动子下游的外源基因得到高水平表达。1993年,毕赤酵母表达系统被Phillip Petrocleum公司出售给RCT(Research Corporation Technologies),在RCT控制下的Invitrogen公司被授权可以向世界范围出售这一系统。目前,常见的表达载体有胞内表达型载体pHIL-D2、pPICZ、pHWO10、pAO815、pGAPZ、pPIC3K,分泌表达型载体pPIC9K、pHIL-S1、pPICZa、pGAPZα等。不同载体具有各自特殊的基因组件,但也有一些共同的序列。如这些载体都包含由醇氧化酶-1(alcohol oxidase 1)基因(AOX1)的启动子和转录终止子(5'AOX1和3'AOX1)组成的表达盒,另外还有一个多克隆位点。外源基因可以在多克隆位点(multiple cloning site,MCS)插入,有些载体还包含组醇脱氢酶(histidinol dehydrogenase)基因(HIS4)选择标记,同时还含有在细菌中复制所必需的序列及选择标记,如大肠杆菌复制起始位点和Ampr基因等。pGAP载体的GAP启动子是自主表达的启动子,不需要甲醇诱导。整合型载体的5'AOX1和3'AOX1可与宿主染色体上的同源基因重组,其结果是载体和外源基因同时插入宿主染色体上,外源基因在5'AOX1启动子控制下表达,其表达水平可占总蛋白的35%以上。
Easyselect pichia表达载体是一种高效表达载体,该载体含有的Zeocin抗性基因可用于阳性重组子的快速筛选,“Easyselect”即为容易筛选的意思。在有些情况下,Easyselect pichia表达载体可以产生多个拷贝,这种重组酵母的产生与酵母菌转化方式有关,概率很小。其载体特征如图8-5所示。该载体具有5'AOX1启动子,可以分为两种:一种是无α因子的,如载体pPICZA、B、C;另一种是含有α因子的,如pPICZaA、B、C,该类载体的终止密码子处有6个组氨基酸序列,形成的表达产物在纯化时可与亲和层析柱结合的镍螯合,便于分离。(https://www.daowen.com)
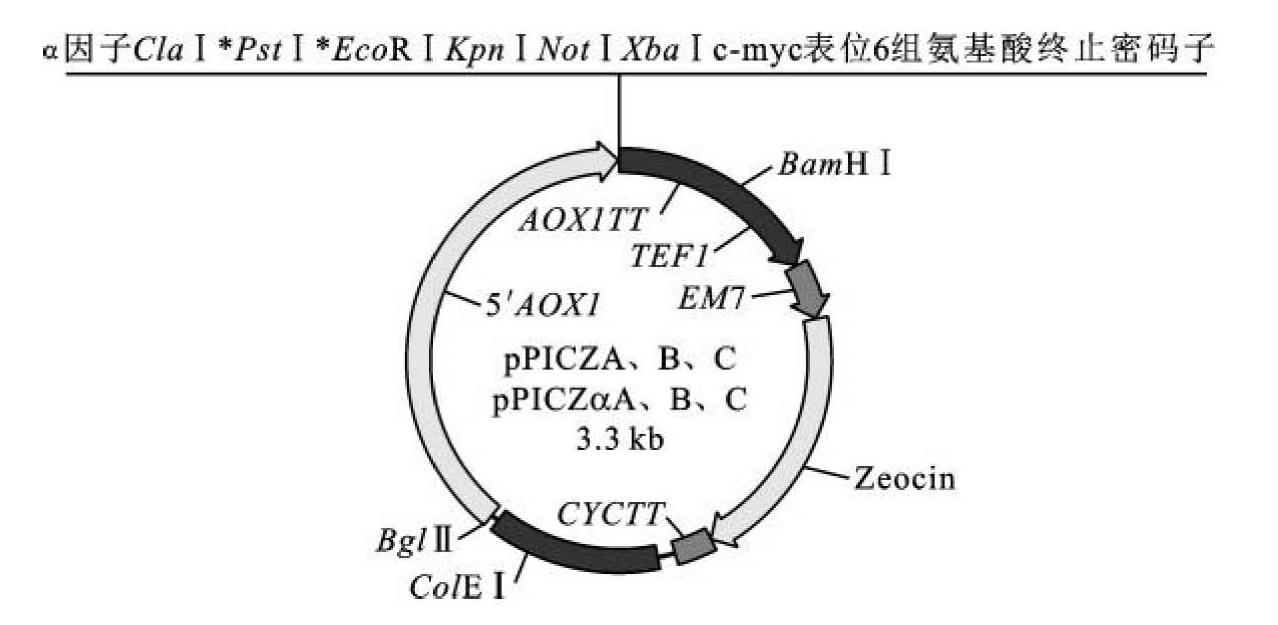
图8-5 Easyselect pichia表达载体
(引自童光志等,2009)
Multi-copy pichia表达载体是一种用于多拷贝目的基因的载体,插入酵母基因组的外源基因以多拷贝形式存在,因此外源基因的表达水平较高。“Multi-copy”就是多拷贝的意思,这类载体有pPIC9K、pPIC3.5K、pAO815三种,其特征如图8-6所示。pPIC9K和pPIC3.5K除了多克隆位点有微小差异外,其酶切位点和抗性筛选标记基本相同,其所含的Kanr基因用于转化子筛选。pPIC3.5K在多克隆位点比pPIC9K多一个BamH Ⅰ位点。载体pPIC9K带有Sig,编码α因子N端信号肽序列,可引导蛋白分泌。而pAO815与它们的结构差异要大一些,没有Kanr基因。pAO815在5'-AOX1-MCS-TT的两端为Bgl Ⅱ和BamH Ⅰ位点,Bgl Ⅱ和BamH Ⅰ为同尾酶,酶切片段可以相互连接,可用于构建多拷贝的目的基因,实现目的基因多拷贝表达。
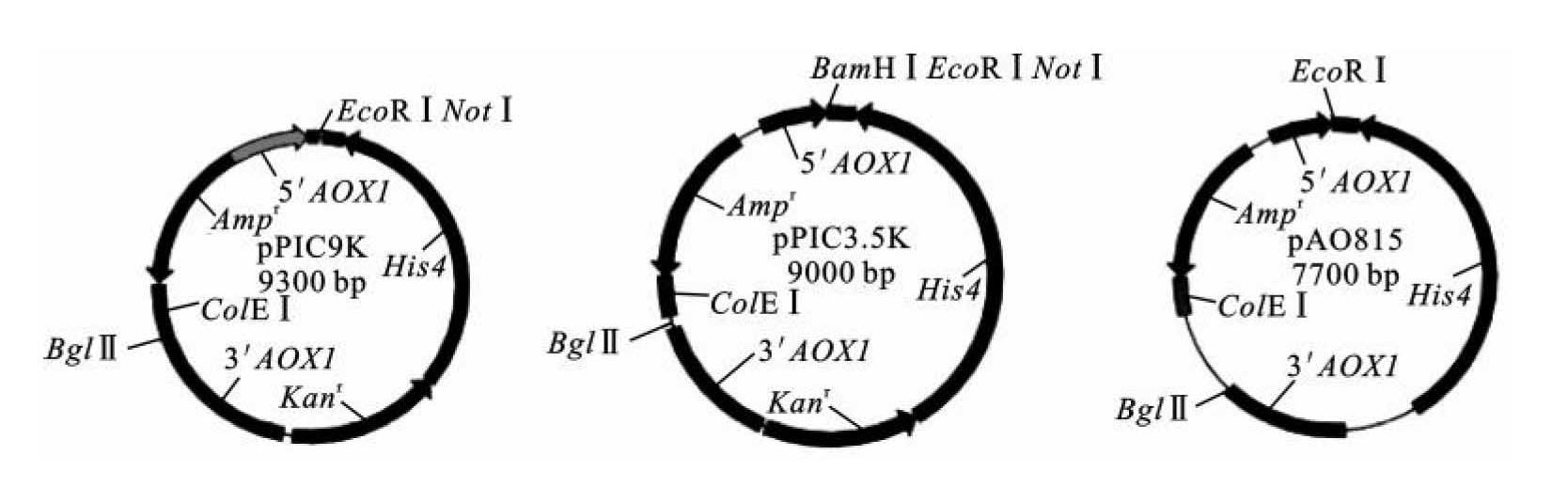
图8-6 Multi-copy pichia表达载体
(引自童光志等,2009)
3.酵母人工染色体
为了克隆更大的外源DNA片段,利用酵母人工染色体(yeast artificial chromosome,YAC)是一种理想的克隆策略。1983年,Murray等首次构建出55 kb酵母人工染色体,它由酵母染色体的DNA复制起始序列(ARS,自主复制序列)、着丝粒、端粒以及酵母选择性标记组成,能自我复制。两端有四膜虫的端粒(TEL,telomere)来保护YAC,其左臂含有酵母筛选标记TRP1、自主复制序列ARS及着丝粒CEN,其右臂含有酵母筛选标记URA3。在两者之间插入外源DNA大片段。酵母人工染色体可插入100~2000 kb的外源DNA片段。由于YAC可容纳更大的DNA片段,因此用不多的克隆就可以包含特定的基因组全部序列,使其迅速成为真核基因组物理图谱制作,以及致病基因克隆和分离的一个重要工具。YAC载体的克隆程序如图8-7所示。
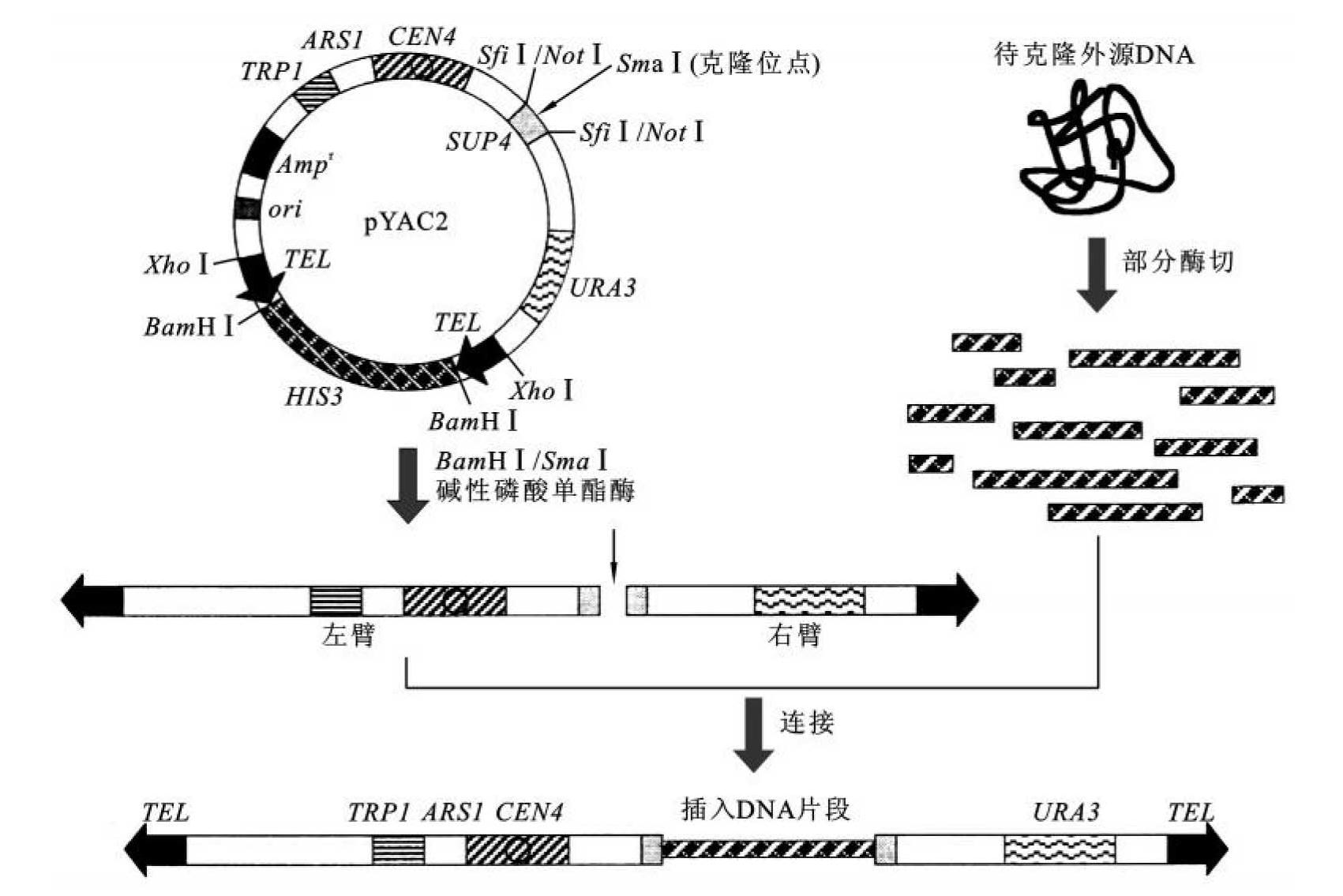
图8-7 酵母YAC载体的克隆程序
(引自张惠展,2005)
YAC还可用于同源重组改造,直接转化哺乳动物细胞,研究基因表达和调节。
常见的YAC克隆载体有三种:pYAC3、pYAC4和pYAC5。3个载体的SUP4基因位置不同,分别在Sna Ⅰ、EcoR Ⅰ、Not Ⅰ位点。以pYAC4为例,该载体含有酵母自主复制序列(ARS1)、着丝粒元件(CEN4)和端粒序列(TEL)。此外生长选择标记URA3可用于选择细胞中的YAC。用含ADE2-1赭石突变的酵母来选择重组子,ADE2-1赭石突变受SUP4基因产物抑制,当外源DNA插入使SUP4基因失活时,形成的菌落为红色。