5.4.1 cDNA文库的构建
基因组含有的全部基因在特定的组织细胞中只有部分表达,处在不同的环境条件和不同分化时期的细胞,其基因表达的种类和强度也不同,所以cDNA文库具有组织细胞特异性。构建cDNA文库通常包括下列步骤。
1.mRNA提取
根据研究目标合理选择组织细胞,利用poly(A)尾巴纯化或直接提取mRNA。常用的寡聚(dT)纤维素亲和层析柱,可以Oligo(dT)吸附mRNA的poly(A)尾,其他不具poly(A)尾的RNA自柱中流出,然后将mRNA洗脱下来。
2.cDNA合成
由mRNA到cDNA的过程称为逆转录,常用禽类成髓细胞病毒(AMV)逆转录酶和鼠白血病病毒(MMLV)逆转录酶催化。
(1)cDNA第一链的合成。
逆转录酶是依赖RNA的DNA聚合酶,合成DNA需要引物引导。常用引物如下。
①Oligo(dT)引物:常用Oligo(dT)引物与mRNA 3'末端的poly(A)配对,引导逆转录酶以mRNA为模板合成第一链cDNA(图5-8)。
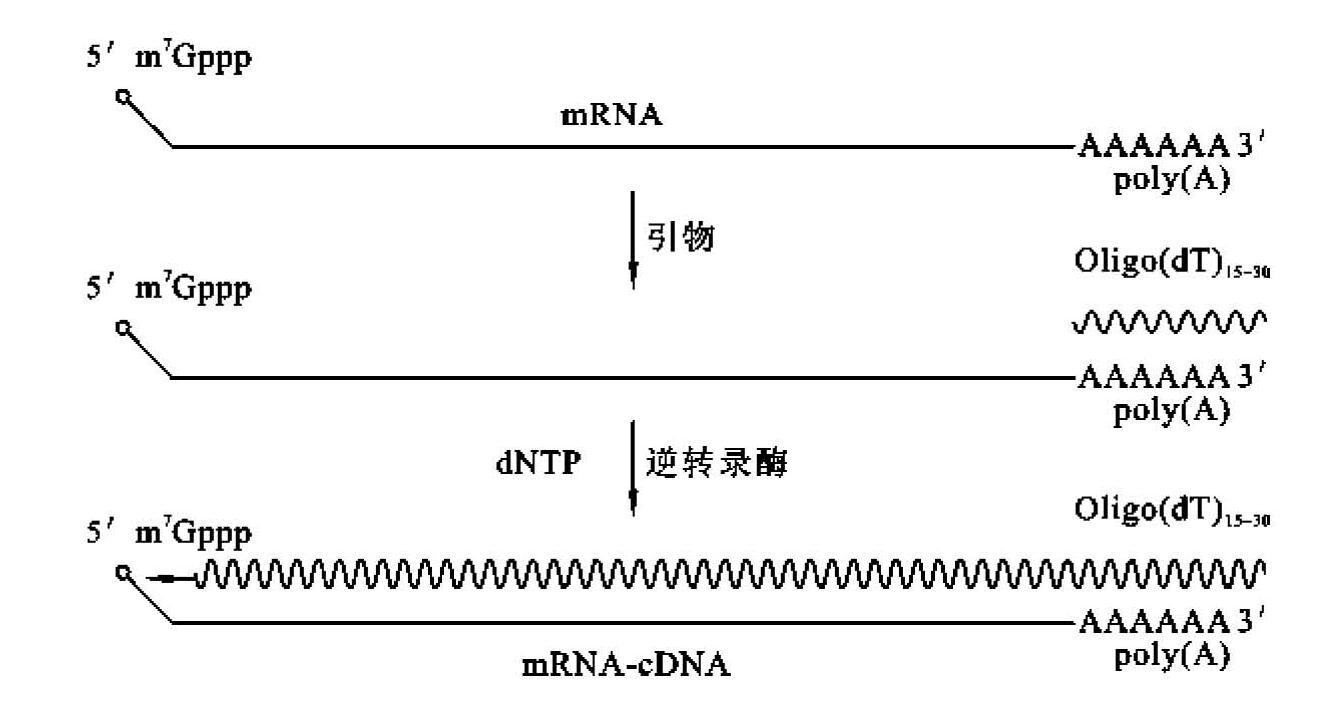
图5-8 逆转录合成单链cDNA
②随机引物:常用化学合成的6~10 nt随机引物,与mRNA分子多处配对,从多个位置合成cDNA第一链,能产生近全长cDNA,适合得到cDNA的5'末端。
③基因特异引物(gene-specific primer,GSP):仅逆转录特定目的基因cDNA的5'末端。
(2)cDNA第二链的合成。(https://www.daowen.com)
cDNA第二链的合成是将mRNA-cDNA杂交双链变成互补双链cDNA。cDNA第二链的合成有以下四种方法。
①自身引导法:合成的单链cDNA 3'末端能形成一个发夹结构,为第二链的合成提供引物,引导第二链的合成。当第一链合成反应产物的DNA-RNA杂交链变性后,利用大肠杆菌DNA聚合酶Ⅰ或逆转录酶合成cDNA第二链,最后用对单链特异性的S1核酸酶消化该环处理去除发夹结构,即可进一步克隆。缺点:在以S1核酸酶切割cDNA的发夹结构时,会导致对应于mRNA 5'末端的序列出现缺失和重排,得到的5'末端不完整,且S1核酸酶处理不好控制,易导致cDNA的丢失,所以该法现已少用。
②置换合成法:第一链在逆转录酶作用下产生的cDNA-mRNA杂交链,不用碱变性,而是在dNTP存在下,利用RNase H在杂交体mRNA链上造成切口和缺口,产生一系列mRNA短片段,作为合成第二链的引物,在大肠杆菌DNA聚合酶Ⅰ和DNA连接酶作用下合成cDNA第二链(图5-9)。此法可得到近全长cDNA,但5'末端可能缺失短序列。置换合成法的优点:合成cDNA效率高;直接利用第一链反应产物,无须进一步处理和纯化;不必使用S1核酸酶来切割双链cDNA中的单链发夹环。
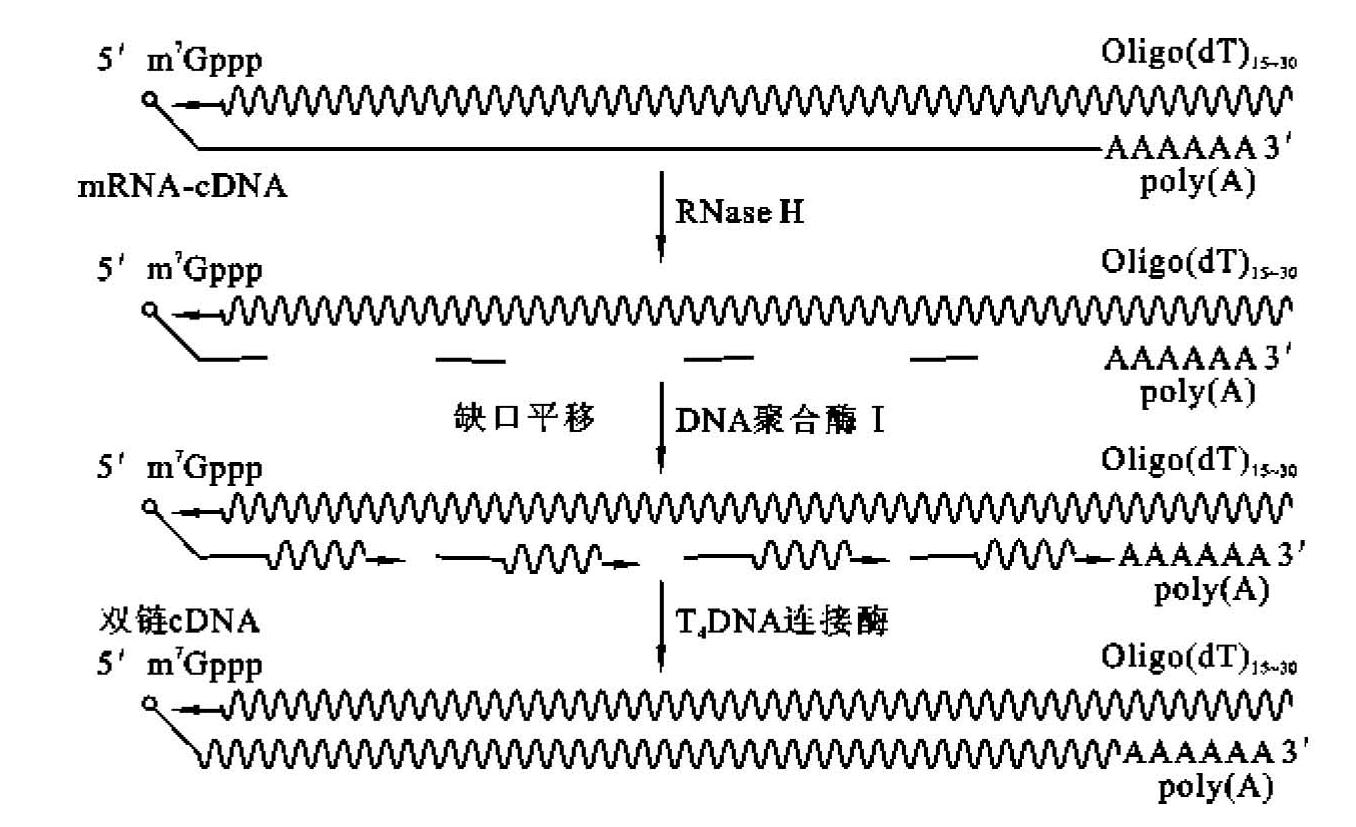
图5-9 置换合成法合成cDNA第二链
③引导合成法:利用末端转移酶在cDNA第一链的3'末端加上poly(dC)尾巴,利用poly(dG)为引物引导第二链合成,利用同源多聚尾与载体进行重组。
④引物-衔接头合成法:此法是引导合成法的改进型,如图5-10所示,在逆转录引物和poly(dG)引物的5'末端引入人工接头,可用PCR扩增总cDNA,也为总cDNA与载体的连接提供酶切位点。
3.双链cDNA的分子克隆
双链cDNA合成后,用末端转移酶给双链分子加尾,或者将人工合成的衔接物加到双链cDNA两端,再同经适当处理而具有相应末端的质粒或噬菌体载体连接。
4.双链cDNA克隆进质粒或噬菌体载体并导入宿主中繁殖
将构成的重组体分子导入大肠杆菌宿主细胞进行扩增,便得到所需的cDNA文库。
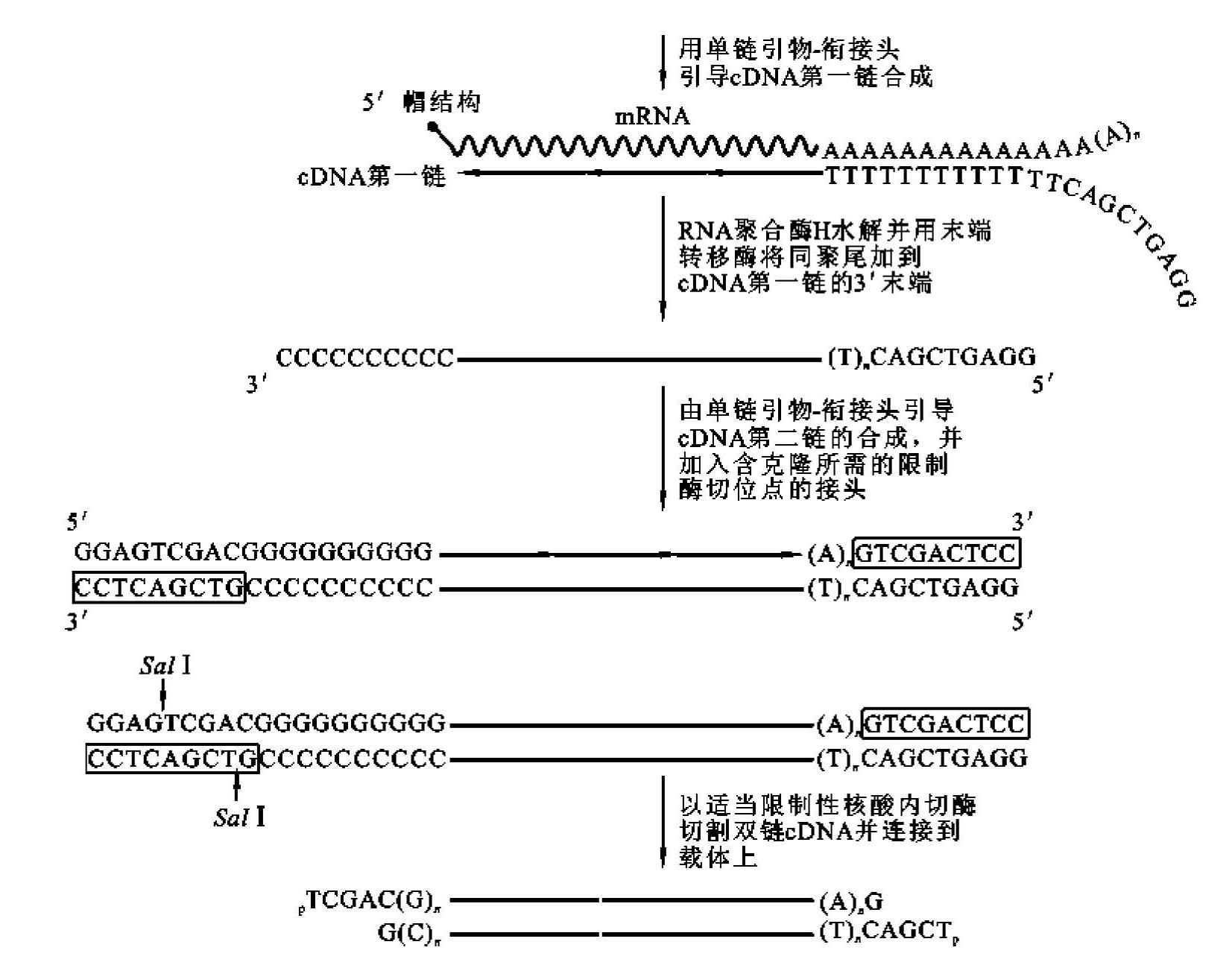
图5-10 引物-衔接头法合成cDNA第二链